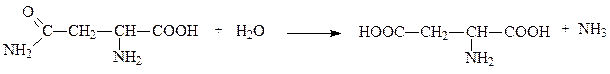
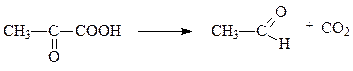
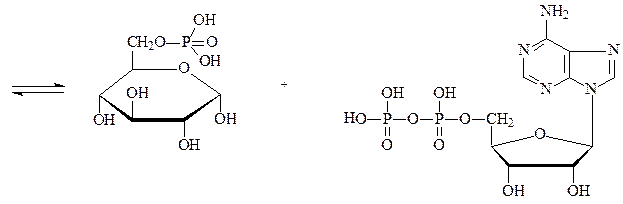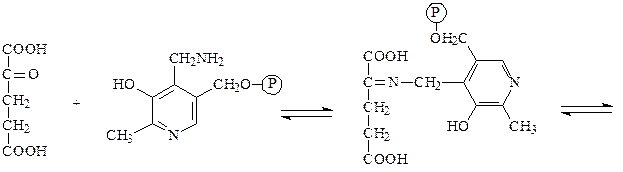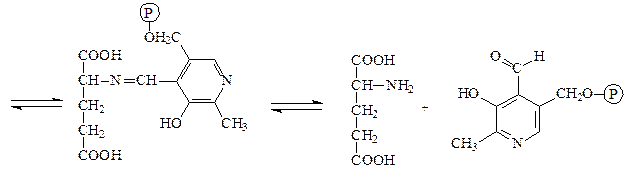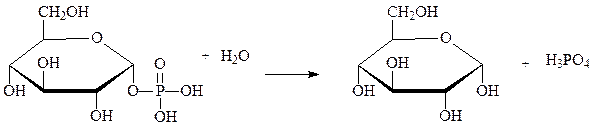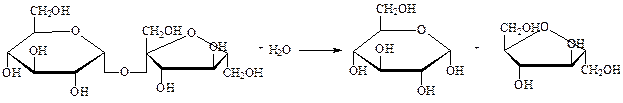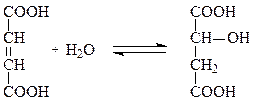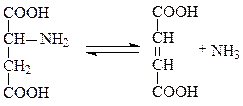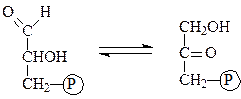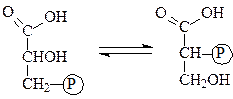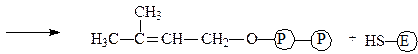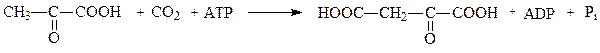Задание 1. Нужно написать уравнения реакции с участием ферментов: липаза, орнитиндекарбоксилаза, амилаза, глюкозофосфатизомераза. Указать класс и подкласс ферманта. Решение:
1.Липаза
Липаза (lipase) — фермент класса гидролаз, катализирующий гидролиз сложноэфирных связей в триглицеридах с образованием жирной кислоты и глицерина. Липазы – ферменты, которые контролируют гидролиз нейтральных жиров (триацилглицеролов):
Класс 3. Гидролазы; Подкласс 3.1. Действующие на сложные эфиры. Подкласс 3.1. Действующие на сложные эфиры. Подподкласс 3.1.1. Гидролазы карбоновых кислот. Классификационный номер — КФ 3.1.1.3.
Характеристика фермента: Систематическое название — Триацилглицерол:ацилгидролаза. Рабочее название — ТАГ-липаза
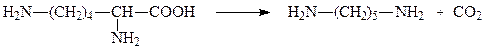
2. Орнитиндекарбоксилаза
Орнитиндекарбоксилаза — высокоспецифичный фермент, катализирует превращение орнитина в путресцин:
Путресцин (диаминобутан) является трупным ядом.
Орнитиндекарбоксилаза относится к классу лиазы, подклассу — углерод-углерод-лиазы. Лиазы – ферменты, катализирующие разрыв С—О, С—С, C—N и других связей, а также обратимые реакции отщепления различных групп негидролитическим путем.
Классификация орнитиндекарбоксилаз: Класс 4. Лиазы; Подкласс 4.1. Углерод-углерод-лиазы.
3. Амилаза
Амилаза — фермент, который катализирует реакцию гидролиза крахмала, гликогена и родственных поли- и олигосахаридов. Амилаза, присутствующий в слюне, катализирует выделение глюкозы из крахмала. При оптимальной температуре и pH одна молекула амилазы может производить 18 ООО молекул глюкозы в секунду.
Классификация: Класс 3. Гидролазы, катализирующие гидролиз химических связей. Подкласс 3.2. Действующие на сахар (углевод).
4. Глюкозофосфатизомераза.
Глюкозофосфатизомераза относится к эпимиразам. Эпимеразы изменяют конфигурацию при одном из хиральных атомов углерода, например: взаимопревращение ?- и ?-изомеров, превращения рибулоза — ксилулоза, галактоза — глюкоза, манноза — галактоза.
Глюкозофосфатизомераза — катализирует взаимное превращение глюкозо-6-фосфата и фруктозо-6-фосфата:
Классификация: Класс 5. Изомеразы; Подкласс 5.1. Рацемазы и эпимеразы.
Задание 2. Помогите написать уравнения реакции с участием ферментов имеющих следующий шифр: КФ 1.1.1.27., КФ 3.1.1.3., КФ 3.1.2.7., КФ 2.7.1.1. Решение:
1. КФ 1.1.1.27.
Лактатдегидрогеназа (L-лактат: NAD-оксидоредуктаза (LDH) 1.1.1.27.) — фермент, принимающий участие в реакциях гликолиза. Лактатдегидрогеназа катализирует превращение лактата в пируват, при этом образуется NADH.
Например, катализирует следующую реакцию:
Лактат + НАД Лактатдегидрогеназа → Пируват + НАДН
ЛДГ катализирует передачу восстановительного эквивалента от лактата на НАД+ (NАD+) или от НАДН (NADH) на пируват.
К.Ф.3.1.1.3. — это фермент липаза или триацилглицерол-липаза широко распространена в природе и играет важную роль в процессах, протекающих при переработке и хранении пищевых продуктов. В настоящие время выделены и охарактеризованы липазы растительного происхождения (липаза клещевины, пшеницы и других злаков), животного (панкреатическая липаза, липаза молока) и микробного (бактериальные и грибные липазы).
Липазы являются универсальными ферментами, используемыми для преобразования липидов. Субстратами липаз являются глицериды и другие сложные эфиры. Липазы более других представителей класса гидролаз обладают способностью катализировать различные типы реакций. Эти типы следующие:
1 – Гидролиз эфиров:
2 – Синтез эфиров:
Обычно липазы катализируют реакцию расцепления триглицеридов согласно приведенному ниже уравнени:
3. КФ 3.1.2.7.
КФ 3.1.2.7. — это глутатионтиолестераза (S- ацилглутатион гидролаза) или (цитрил-глутатион тиоэфиргидролаза). Этот фермент катализирует следующую химическую реакцию:
S- ацилглутатион + H2O → глутатион + карбоксилат
3. КФ 3.1.2.7.
КФ 2.7.1.1. — это фермент гексокиназа (ATР: D-гексозо-6-фосфотрансфераза), катализирует перенос фосфатной группы с АТР на моносахариды:
Ферменты. Химическая природа, физико-химические свойства и биологическая роль.
Основу жизнедеятельности любого организма составляют химические процессы. Практически все реакции в живом организме протекают с участием природных биокатализаторов, называемых ферментами или энзимами. Ферменты – это белки (установлено в 1922г), которые действуют как катализаторы в биологических системах.
Являясь веществами белкой природы, ферменты обладают всеми свойствами белков:
являются амфотерными соединениями;
вступают в те же качественные реакции, что и белки (биуретовую, ксантопротеиновую, фолина и др.);
подобно белкам растворяются в воде с образованием коллоидных растворов;
обладают электрофоретической активностью;
гидролизуются до аминокислот;
склонны к денатурации под влиянием тех же факторов: температуры, изменениях рН, действием солей тяжелых металлов, действием физических факторов (ультразвук, ионизирующее излучение и др.);
имеют несколько уровней организации макромолекул, что подтверждено данными рентгеноструктурного анализа, ЯМР, ЭПР.
Биологическая роль ферментов заключается в том, что они катализируют контролируемое протекание всех метаболических процессов в организме.
Сравнение каталитического действия ферментов и неорганических катализаторов
Сходство ферментов инеорганических катализаторов
Отличие ферментов отнеорганических катализаторов
1. Ускоряют только термодинамически возможные реакции
1. Для ферментов характерна высокая специфичность: субстратная специфичность:
абсолютная (1 фермент – 1 субстрат),
групповая (1 фермент – несколько похожих субстратов)
стереоспецифичность (ферменты работают с субстратами только определенного стереоряда L или D ).
каталитическая специфичность (ферменты катализируют реакции преимущественно одного из типов химических реакций – гидролиза, окисления-восстановления и др)
2. Не изменяют состояние равновесия реакций, а только ускоряют его достижение.
2. Высокая эффективность действия: ферменты ускоряют реакции в10 8 -10 14 раз.
3. В реакциях не расходуются
3. Ферменты действуют только в мягких условиях ( t = 36-37ºС, рН
7,4, атмосферное давление), т.к. они обладают конформационной лабильностью – способностью к изменению конформации молекулы под действием денатурирующих агентов (рН, Т, химические вещества).
4. Действуют в малых количествах
4. В организме действие ферментов регулируется специфически (катализаторы только неспецифически)
5. Чувствительны к активаторам и ингибиторам
5. Широкий диапазон действия (большинство процессов в организме катализируют ферменты).
В настоящее время учение о ферментах является центральным в биохимии и выделено в самостоятельную науку – энзимологию. Достижения энзимологии используются в медицине для диагностики и лечения, для изучения механизмов патологии, а, кроме того, и в других областях, например, в сельском хозяйстве, пищевой промышленности, химической, фармацевтической и др.
Классификация и номенклатура ферментов
Классификация ферментов основана на типе катализируемой химической реакции; На основании 6 типов химических реакций ферменты, которые их катализируют, подразделяют на 6 классов, в каждом из которых несколько подклассов и поподклассов (4-13); Каждый фермент имеет свой шифр КФ 1.1.1.1. Первая цифра обозначает класс, вторая – подкласс, третья – подподкласс, четвертая – порядковый номер фермента в его подподклассе (в порядке открытия). Название фермента состоит из 2 частей: 1 часть – название субстрата (субстратов), 2 часть – тип катализируемой реакции. Окончание – АЗА; Дополнительная информация, если необходима, пишется в конце и заключается в скобки: L-малат + НАДФ+ ↔ ПВК + СО2 + НАДН2 L-малат: НАДФ+ – оксидоредуктаза (декарбоксилирующая); В правилах названия ферментов нет единого подхода.
1. Оксидоредуктазы Катализируют окислительно-восстановительные реакции. В реакцию вступают 2 вещества и 2 образуются, одно окисляется, другое восстанавливается: Sвост + S’окисл ↔ S’вост + Sокисл Оксидоредуктазы делятся на: дегидрогеназы (отщепляют Н от субстратов), оксидазы (переносят Н с субстрата на кислород), оксигеназы (включают кислород в молекулу субстрата), гидроксипероксидазы (разрушают перекиси водорода и органические перекиси). Систематическое название включает в себя название донора е и Н+ через двоеточие название акцептора через тире – название класса: донор: акцептор ( косубстрат) оксидоредуктаза
2. Трансферазы Ферменты этого класса принимают участие в переносе атомных групп, молекулярных остатков от одного соединения к другому. В реакцию вступают 2 вещества и 2 образуются: S-G + S’ ↔ S + S’-G. В зависимости от переносимых групп трансферазы делятся на: 1). фосфотрансферазы (киназы); 2). аминотрансферазы; 3). гликозилтрансферазы; 4). метилтрансферазы; 5). ацилтрансферазы. Систематическое название: откуда: куда в какое положение–что–трансфераза или донор: акцептор–транспортируемая группа– трансфераза
3. Гидролазы. Расщепляют ковалентную связь с присоединением молекулы воды. В реакцию вступают 2 вещества и 2 образуются: S-G + Н2О ↔ S-ОН + G-Н. В зависимости от характера гидролизуемой связи, различают подклассы: 1). гликозидазы – гидролиз гликозидов (лактоза – лактаза, мальтоза – мальтаза, сахароза – сахараза); 2). пептидазы – гидролиз пептидных связей; 3). эстеразы – разрыв связи в сложных эфирах. Систематическое название субстрат–что отщепляется–гидролаза или субстрат–гидролаза Ацетилхолин + Н2О ↔ Ацетат + Холин Систематическое название: Ацетилхолин-ацилгидролаза (Ацетилхолин-гидролаза) Тривиальное название: Ацетилхолинэстераза
4. Лиазы Отщепление групп от субстратов по негидролитическому механизму с образованием двойных связей (или наоборот, присоединение по двойной связи). Реакции обратимы, за исключением отщепления СО2. В реакцию вступает 1 вещество и 2 образуются (или наоборот): -SХ-SY- ↔ XY + -S=S- Систематическое название субстрат: что отщепляется–лиаза L-малат ↔ фумарат + Н2О Систематическое название: L-малат: гидро–лиаза Тривиальное название: фумараза
5. Изомеразы Взаимопревращения оптических, геометрических, позиционных изомеров. В реакцию вступает 1 вещество и 1 образуется. Исходя из типа катализируемой реакции изомеризации выделяется несколько подклассов:
рацемазы;
эпимеразы;
таутамеразы;
цис,- трансизомеразы;
мутазы (при внутримолекулярном переносе группы);
цикло-, оксоизомеразы.
Систематическое название субстрат–вид изомеризации–изомераза или субстрат–продукт–изомераза Фумаровая к-та ↔ малеиновая к-та Систематическое название: фумарат–цис,транс–изомераза
Ферменты – это специфические белки, которые действуют как катализаторы в биологических системах. Являясь веществами белкой природы, ферменты обладают всеми свойствами белков:
гидролизуются до аминокислот
дают положительные цветные реакции на белки (биуретовую, ксантопротеиновую).
подобно всем белкам растворяются в воде с образованием коллоидных растворов.
являются амфотерными соединениями.
склонны к денатурации под влиянием тех же факторов: температуры, изменениях рН, действием солей тяжелых металлов, действием физических факторов (ультразвук, ионизирующее излучение и др.).
наличие нескольких уровней организации макромолекул, подтвержденное данными ренгеноструктурного анализа. В отличии от других белков ферменты обладают каталитическими свойствами. При этом, обладая свойствами, присущими неорганическим катализаторам, белки существенно отличаются от них рядом свойств (табл.1).
Таблица 1. Сходство и отличие каталитического действия ферментов и неорганических катализаторов.
1. Ускоряют только термодинамически возможные реакции
2. В случае необратимых реакций, ускоряют и прямую и обратную реакцию
3. Чувствительны к активаторам и ингибиторам
4. Действуют в малых количествах
1. Действуют в мягких условиях ( t = 36-37ºС, рН
7,4, атмосферное давление)
2. Действие зависит от рН среды
4. Высокая специфичность
5. Действие ферментов в организме регулируется
6. Широкий диапазон ферментативного действия.
7. Высокая эффективность действия.
Строение ферментов
Метаболит – вещество, которое участвует в метаболических процессах. Субстрат – вещество, которое вступает в химическую реакцию. Продукт – вещество, которое образуется в ходе химической реакции. Ферменты характеризуются наличием специфических центров катализа.
Активный центр (Ац) – это часть молекулы фермента, которая специфически взаимодействует с субстратом и принимает непосредственное участие в катализе. Ац, как правило, находиться в нише (кармане). В Ац можно выделить два участка: участок связывания субстрата – субстратный участок (контактная площадка) и собственно каталитический центр.
Большинство субстратов образует, по меньшей мере, три связи с ферментом, благодаря чему молекула субстрата присоединяется к активному центру единственно возможным способом, что обеспечивает субстратную специфичность фермента. Каталитический центр обеспечивает выбор пути химического превращения и каталитическую специфичность фермента. У группы регуляторных ферментов есть аллостерические центры, которые находятся за пределами активного центра. К аллостерическому центру могут присоединяться “+” или “–“ модуляторы, регулирующие активность ферментов.
Различают ферменты простые, состоят только из аминокислот, и сложные, включают также низкомолекулярные органические соединения небелковой природы (коферменты) и (или) ионы металлов (кофакторы).
Коферменты – это органические вещества небелковой природы, принимающие участие в катализе в составе каталитического участка активного центра. В этом случае белковую составляющую называют апоферментом, а каталитически активную форму сложного белка – холоферментом. Таким образом: холофермент = апофермент + кофермент.
В качестве коферментов функционируют:
гемы,
нуклеотиды,
коэнзим Q,
ФАФС,
SAM,
Глутатион
производные водорастворимых витаминов:
РР (никотиновая кислота)
Кофермент, который присоединен к белковой части ковалентными связями называется простетической группой. Это, например, FAD, FMN, биотин, липоевая кислота. Простетическая группа не отделяется от белковой части. Кофермент, который присоединен к белковой части нековалентными связями называется косубстрат. Это, например, НАД+, НАДФ+. Косубстрат присоединяется к ферменту в момент реакции.
Кофакторы ферментов – это ионы металлов, необходимые для проявления каталитической активности многих ферментов. В качестве кофакторов выступают ионы калия, магния, кальция, цинка, меди, железа и т.д. Их роль разнообразна, они стабилизируют молекулы субстрата, активный центр фермента, его третичную и четвертичную структуру, обеспечивают связывание субстрата и катализ. Например, АТФ присоединяется к киназам только вместе с Mg2+.
Изоферменты – это множественные формы одного фермента, катализирующие одну и ту же реакцию, но отличающие по физическим и химическим свойствам (сродству к субстрату, максимальной скорости катализируемой реакции, электрофоретической подвижности, разной чувствительности к ингибиторам и активаторам, оптимуму рН и термостабильности). Изоферменты имеют четвертичную структуру, которая образована четным количеством субъединиц (2, 4, 6 и т.д.). Изоформы фермента образуются в результате различных комбинаций субъединиц.
Можно выделить три группы коферментов: 1. Принимающие участие в окислительно-восстановительных реакциях: НАД, ФАД, KoQ. 2. Принимающие участие в переносе атомных групп: фосфопиридоксаль (активная форма В6, перенос аминогрупп), биотин (перенос диоксида углерода), перенос метильных и других групп. 3. Принимающие участие в реакциях синтеза, изомеризации и расщепления: тиаминпирофосфат (активная форма В1, декарбоксилирование кетокислот), тиаминдифосфат, коэнзим А (для активации субстрата, например – жирные кислоты).
В качестве примера можно рассмотреть лактатдегидрогеназу (ЛДГ), фермент, который катализирует обратимую реакцию:
ЛДГ существует в виде 5 изоформ, каждая из которых состоит из 4-х протомеров (субъединиц) 2 типов М (muscle) и Н (heart). Синтез протомеров М и Н типа кодируется двумя разными генетическими локусами. Изоферменты ЛДГ различаются на уровне четвертичной структуры: ЛДГ1 (НННН), ЛДГ2 (НННМ), ЛДГ3 (ННММ), ЛДГ4 (НМММ), ЛДГ5 (ММММ). Полипептидные цепи Н и М типа имеют одинаковую молекулярную массу, но в составе первых преобладают карбоновые аминокислоты, последних – диаминокислоты, поэтому они несут разный заряд и могут быть разделены методом электрофореза.
Кислородный обмен в тканях влияет на изоферментный состав ЛДГ. Где доминирует аэробный обмен, там преобладают ЛДГ1, ЛДГ2 (миокард, надпочечники), где анаэробный обмен – ЛДГ4, ЛДГ5 (скелетная мускулатура, печень). В процессе индивидуального развития организма в тканях происходит изменение содержания кислорода и изоформ ЛДГ. У зародыша преобладают ЛДГ4, ЛДГ5. После рождения в некоторых тканях происходит увеличение содержания ЛДГ1, ЛДГ2. Существование изоформ повышает адаптационную возможность тканей, органов, организма в целом к меняющимся условиям. По изменению изоферментного состава оценивают метаболическое состояние органов и тканей.
Локализация и компартментализация ферментов в клетке и тканях. Ферменты по локализации делят на 3 группы: I – общие ферменты (универсальные) II – органоспецифические III – органеллоспецифические
Общие ферменты обнаруживаются практически во всех клетках, обеспечивают жизнедеятельность клетки, катализируя реакции биосинтеза белка и нуклеиновых кислот, образование биомембран и основных клеточных органелл, энергообмен. Общие ферменты разных тканей и органов, тем не менее, отличаются по активности.
Органоспецифичные ферменты свойственны только определенному органу или ткани. Например: Для печени – аргиназа. Для почек и костной ткани – щелочная фосфатаза. Для предстательной железы – КФ (кислая фосфатаза). Для поджелудочной железы – α-амилаза, липаза. Для миокарда – КФК (креатинфосфокиназа), ЛДГ, АсТ и т.д. Внутри клеток ферменты также распределены неравномерно. Одни ферменты находятся в коллоидно-растворенном состоянии в цитозоле, другие вмонтированы в клеточных органеллах (структурированное состояние).
Органеллоспецифические ферменты. Разным органеллам присущ специфический набор ферментов, который определяет их функции. Органеллоспецифические ферменты это маркеры внутриклеточных образований, органелл:
Ядро клетки: ферменты обеспечивающие синтез РНК, ДНК ( РНК-полимераза, НАД-синтетаза).
Ядрышко: ДНК-зависимая-РНК-полимераза
В результате в клетке образуются отсеки (компартменты), которые отличаются набором ферментов и метаболизмом (компартментализация метаболизма).
Регуляторные ферменты – это относительно немногочисленная группа ферментов, которые способны отвечать на специфические регуляторные воздействия изменением активности. Эти ферменты локализуются в начале или в местах разветвления метаболических путей.
Среди ферментов выделяется немногочисленная группа регуляторных ферментов, которые способны отвечать на специфические регуляторные воздействия изменением активности. Эти ферменты имеются во всех органах и тканях и локализуются в начале или в местах разветвления метаболических путей.Строгая локализация всех ферментов закодирована в генах. Определение в плазме или сыворотке крови активности органо- органеллоспецифических ферментов широко используется в клинической диагностике.
Увеличение активности ферментов в плазме (сыворотке) крови связано прежде всего с цитолизом (т.е. повышением проницаемости биомембран или их некрозом) и выходом ферментов в кровяное русло. При этом активность ферментов в поврежденном органе уменьшается, а в плазме (сыворотке) крови возрастает. По измерению активности органеллоспецифических ферментов можно судить о тяжести патологического процесса.
Единицей ферментативной активности, рекомендуемой для употребления международным биохимическим союзом, является международная единица активности (МЕ).
МЕ – это количество фермента, которое катализирует превращение 1 мкМоля субстрата за 1 минуту (мкМоль/мин) в оптимальных условиях. Однако МЕ не может считатьсяединицей СИ, т.к. «минута» является внесистемной единицей. Тем не менее, в нашей стране, а особенно за рубежом МЕ/л – мкМоль/(мин*л) получила широкое распространение для выражения активности ферментов сыворотки крови. Кроме того, активность ферментов выражалась в различных внеклеточных единицах: мМоль/мин*л, мМоль/ч*л, мкМоль/ч*мл, мМоль/с*л и др. Единицей СИ активности ферментов является катал (кат) и его производные (мккат и др.).
Катал – это количество фермента, которое катализирует превращение 1 Моль субстрата за 1 секунду (Моль/с). Следовательно, активность ферментов в клинико-диагностических исследованиях должна выражаться в каталах и его долях на литр, при этом необходимо помнить, что кат/л = Моль/с*л, мкат/л = мМоль/л*с и т.д. Исключение составляют ферменты, у которых молекулярная масса субстрата неизвестна (например, амилаза). В этих случаях активность можно выражать в единицах массы субстрата, превращенном 1 л сыворотки крови за 1 с или 1 мин.
Ниже приводятся коэффициенты для перевода единиц ферментативной активности:
Триэтаноламиновый буфер, рН 7,2; 25˚С в присутствии тиолов как активаторов
Трис-буфер, рН 7,2; 37˚С, без активаторов
Фосфатный буфер, рН 7,5; 25˚С
Электрофорез в полиакриламидном геле
Фосфатный буфер, рН 7,4; 25˚С
Фосфатный буфер, рН 7,4; 25˚С
Мышьяковистый-HCl-буфер, рН 7,15; 37˚С
Триэтаноламиновый буфер, рН 7,5; 25˚С
Трис-буфер, рН 7,5; 37˚С
Эмульсия триглицеридов (оливковое масло)
Трис-буфер, рН 8.0; 37˚С
Фосфатный буфер 115 М, рН 7,8; 37˚С
Барбиталовый буфер, рН 5,0; 37˚С
Цитратный буфер, рН 4,8; 37˚С
Цитратный буфер в присутствии солей винной кислоты, рН 4,8; 37˚С
Барбиталовый буфер, рН 9,6; 37˚С
Глициновый буфер, рН 10,4; 37˚С
Гидразиновый буфер, рН 7,4; 37˚С
Коллидиновый буфер, рН 7,4; 37˚С
Коллидиновый буфер, рН 7,4; 37˚С
Коллидиновый буфер, рН 7,4; 37˚С
Фосфатный буфер, рН 7,2; 25˚С
Фосфатный буфер, рН 7,7; 25˚С
Примечание. Необходимо подчеркнуть, что получаемые величины активности можно трактовать только с учетом используемого метода и конкретных условий определения.
Регуляция активности ферментов в клетке. Общие представления о гормонах и их роли в регуляции активности ферментов.
Одним из важнейших свойств живых организмов является способность к поддержанию гомеостаза. Гомеостаз в организме поддерживается за счет регуляции скорости ферментативных реакций.
Эта регуляция осуществляется:
Доступностью молекул субстрата и кофермента;
Изменением каталитической активности молекул фермента;
Изменением количества молекул фермента.
I. Доступность молекул субстратов обеспечивается контролируемой работой трансмембранных транспортных систем. Например, количество в мембране ГЛЮТов (трансмембранных переносчиков глюкозы) определяет скорость поступления глюкозы в цитоплазму клеток и скорость метаболических процессов, в которых она используется (гликолиз, ПФШ, гликогенез).
Доступность коферментов зависит от скорости их регенерации. В результате, чем больше концентрация исходного субстрата и регенерированных коферментов, тем выше скорость метаболического пути. Например, дефицит НАД+ лимитирует реакции ЦТК.
II. Регуляция каталитической активности ферментов. Бывает:
Неспецифическая регуляция. В связи с лабильностью всех ферментов, их каталитическая активность зависит от температуры, рН и давления.
Специфическая регуляция. Под действием специфических активаторов и ингибиторов изменяется активность регуляторных ферментов, которые контролируют интенсивность метаболических процессов в организме.
Механизмы специфической регуляции каталитической активности ферментов:
Аллостерическая регуляция;
Регуляция с помощью белок-белковых взаимодействий;
Регуляция через ковалентную модификацию.
а). Регуляция путем фосфорилирования/дефосфорилирования фермента; б). Регуляция частичным протеолизом.
1). Аллостерическая регуляция каталитической активности ферментов. Аллостерическими ферментами называют ферменты, активность которых регулируется обратимым нековалентным присоединением модулятора (активатора и ингибитора) к аллостерическому центру. Ингибиторами аллостерических ферментов часто являются конечные продукты метаболических путей, активаторами – их начальные субстраты. Активирование происходит по принципу прямой положительной связи, а ингибирование – по принципу отрицательной обратной связи.
Аллостерические ферменты играют важную роль в регуляции т.к. чрезвычайно быстро реагируют на изменения среды. Например, конечный продукт катаболизма глюкозы АТФ ингибирует аллостерически ферменты гликолиза фосфофруктокиназу и пируваткиназу. Накапливаемая в гликолизе фруктоза-1,6-ф активирует пируваткиназу, что ускоряет реакции гликолиза.
2). Регуляция каталитической активности ферментов с помощью белок-белковых взаимодействий.
Выделяют 2 механизма:
Активация ферментов в результате присоединения регуляторных белков. Например, аденилатциклаза (Ац), которая катализирует превращение АТФ в цАМФ, активируется присоединением α-субъединицы G-белка. Этот механизм регуляции обратим.
Регуляция каталитической активности ферментов ассоциацией/диссоциацией протомеров. Например, протеинкиназа А, активируется при диссоциации ее тетрамера на 4 субъединицы и инактивируется при обратном соединении 4 субъединиц в тетрамер.
3). Регуляция каталитической активности ферментов путем их ковалентной модификации. Регуляция активности фермента осуществляется в результате ковалентного присоединения или отщепления от него фрагмента.
Она бывает 2 видов:
а). путем фосфорилирования и дефосфорилирования ферментов;
б). путем их частичного протеолиза.
а). Регуляция каталитической активности ферментов путем их фосфорилирования и дефосфорилирования. Фосфорилирование осуществляется протеинкиназами (ПК) по ОН-группе серина, треонина или тирозина регуляторный белков и ферментов. Дефосфорилирование в этих же положениях осуществляется фосфопротеинфосфатазами (ФПФ).
Введение отрицательно заряженной фосфорной группы приводит к обратимому изменению конформации и активности фермента.
Например, под действием глюкагона и адреналина в клетках печени происходит фосфорилирование ключевых ферментов гликогенеза (гликогенсинтаза) и гликогенолиза (гликогенфосфорилаза), при этом распад гликогена активируется, а синтез ингибируется. Инсулин наоборот вызывает в клетках печени дефосфорилирование тех же ключевых ферментов, в результате синтез гликогена активируется, а распад ингибируется.
б). Регуляция каталитической активности ферментов путем их частичного протеолиза. При участии активаторов и протеолитических ферментов происходит отщепление части молекулы фермента и его необратимая активация. Такой фермент функционирует короткий период, а затем разрушается. Подобная схема активации характерна для внеклеточных ферментов ЖКТ (пепсин, трипсин, химотрипсин и др.) и ферментов свертывающей и противосвертывающей системы крови (тромбин, фибрин, плазмин др.). Например, трипсиноген, синтезируемый в поджелудочной железе, поступает в двенадцатиперстную кишку, где энтеропептидаза кишечника отщепляет у него с N-конца гексапептид. В результате в оставшейся части молекулы фермента формируется активный центр.
III. Механизмы регуляции количества ферментов
Количество ферментов в клетке зависит от скорости их синтеза и распада. Синтез ферментов регулируется индукторами и репрессорами. В качестве индукторов и репрессоров выступают некоторые метаболиты, гормоны и биологически активные вещества.
Индукторы – это вещества которые запускают синтез ферментов. Процесс запуска синтеза ферментов называется индукцией. Например, у бактерий ферменты синтезируются только при наличии для них субстратов, которые являются для этих ферментов индукторами (у E. coli лактоза индуктор β-галактозидазы). Не все ферменты чувствительны к индукторам. Ферменты, концентрация, которых зависит от добавления индукторов, называются индуцируемыми ферментами (органоспецифические ферменты). Ферменты, концентрация которых постоянна и не регулируется индукторами, называются конститутивными ферментами (ферменты гликолиза, синтеза РНК и т.д.). Для индуцируемых ферментов выделяют понятие базовый уровень, это концентрация фермента при отсутствии индуктора. При индукции базовый уровень фермента может быть превышен от 2 до 1000 раз.
Репрессоры (корепрессоры) – вещества, которые останавливают синтез ферментов. Процесс остановки синтеза ферментов называется репрессией.
Дерепрессия – процесс возобновления синтеза ферментов после удаления из среды репрессора или истощения его запасов. Часто один индуктор или репрессор регулируют синтез сразу нескольких ферментов участвующих в одном метаболическом пути. Гены этих ферментов собраны в оперон.
Оперон – участок молекулы ДНК, который содержит информацию о группе функционально взаимосвязанных структурных генов и регуляторную зону, промотор, контролирующую транскрипцию этих генов. Индукция и репрессия синтеза ферментов оперона называется координированной.
Координированная индукция – все ферменты, кодируемые генами оперона, индуцируются одним индуктором.
Координированная репрессия – остановка синтеза ферментов оперона одним репрессором. В промоторе оперона имеется участок оператор, необходимый для присоединения репрессора.
Репрессор синтезируется с гена-регулятора.
Индукция оперона. Репрессор присоединяется к оператору и препятствует присоединению к промотору РНК-полимеразы – транскрипции мРНК нет. Индуктор, присоединяясь к репрессору, вызывает отделение последнего от оператора, РНК-полимераза присоединяется к промотору и начинается транскрипция мРНК с оперона. С этой мРНК синтезируются сразу несколько ферментов.
Репрессия оперона. Репрессор не присоединяется к оператору самостоятельно. Присоединение к оператору репрессора происходит только в комплексе с корепрессором. Комплекс репрессор-корепрессор присоединяясь к оператору, препятствует присоединению РНК-полимеразы к промотору, что блокирует транскрипцию мРНК с оперона и синтез ферментов.
Катаболическая репрессия – более выгодный субстрат окисления ингибирует ферменты окисления менее выгодного субстрата. Распад ферментов идет под действием гидролитических ферментов. Они активно атакуют ферменты, структура которых подверглась модификации (например, денатурации) под действием различных неблагоприятных факторов.
Клеточная сигнализация
В многоклеточных организмах поддержание гомеостаза обеспечивают 3 системы:
Регуляторные системы функционируют с участием сигнальных молекул.
Сигнальные молекулы – это органические вещества, которые переносят информацию. К сигнальным молекулам относятся гормоны, нейромедиаторы, факторы роста, цитокины и эйкозаноиды. ЦНС для передачи сигнала использует нейромедиаторы, гуморальная система – гормоны, иммунная – цитокины.
Гормоны – это сигнальные молекулы беспроводного системного действия. Отличием истинных гормонов от других сигнальных молекул, является то, что они синтезируются в специализированных эндокринных клетках, транспортируются кровью и действуют дистантно на ткани мишени.
Пептидные гормоны и катехоламины растворимы в воде, они регулируют преимущественно каталитическую активность ферментов. Стероидные и тиреоидные гормоны водонерастворимы, они регулируют преимущественно количество ферментов.
Гормоны влияют на активность и количество ферментов в клетке не напрямую, а через каскадные системы (аденилатциклазную, гуанилатциклазную, инозитолтрифосфатную, RAS и т.д.), состоящие из:
рецепторов;
регуляторных белков (G-белки, IRS, Shc, STAT и т.д.).
Необходимость каскадных систем связана с тем, что, во-первых, водорастворимые гормоны не проходят клеточную мембрану, во-вторых, эти системы обеспечивают усиление первичного сигнала гормонов в миллионы раз. В результате даже одна молекула гормона способна активировать миллионы ферментов и вызвать метаболический эффект. Водонерастворимые гормоны самостоятельно проходят клеточные мембраны и реализуют свой эффект с участием цитоплазматических и ядерных рецепторов.
Рецепторы
Рецепторы – это белки, встроенные в клеточную мембрану или находящиеся внутри клетки, которые, взаимодействуя с сигнальными молекулами, меняют активность регуляторных белков.
По локализации рецепторы делятся на:
По эффекту рецепторы делятся на:
активаторные (активируют каскадные системы)
ингибиторные (блокируют каскадные системы).
Участие рецепторов в трансмембранной передаче сигнала:
Рецепторы, связанные с ионными каналами (рецептор ГАМК);
Рецепторы, с тирозинкиназной активностью (рецептор инсулина);
Рецепторы, активирующие инозитолтрифосфатную систему (α1-адренорецептор – у гепатоцитов);
Рецепторы, с гуанилатциклазной активностью (гуанилатциклаза, рецептор ПНФ);
Рецепторы, активирующие аденилатциклазную систему (β-адренорецепторы);
Рецепторы, связывающие гормон в цитозоле или ядре (рецептор кортизола).
По механизму передачи сигнала рецепторы делятся на 4 типа:
Рецепторы, связанные с ионными каналами. Это интегральные мембранные белки, состоящие из нескольких субъединиц, полипептидная цепь которых несколько раз пересекает клеточную мембрану. Они имеют центр связывания сигнальной молекулы и ионный канал. При связывании с сигнальными молекулами у этих рецепторов открываются или закрываются ионные каналы. Действуют они очень быстро в течение миллисекунд. Эти рецепторы обеспечиваю синаптическую передачу в электрически возбудимых клетках. Например, катионные ацетилхолиновые никотиновые рецепторы скелетных мышц.
Рецепторы, с ферментативной активностью. Имеют разнообразное строение, регулируют клеточное деление, дифференцировку, развитие иммунного ответа.
Рецепторы с ферментативной активностью бывают 3 видов:
а). Рецепторы, с тирозинкиназной активностью (тирозиновые протеинкиназы). Это каталитические рецепторы, фосфорилирующие по тирозину белки-мишени. Их активируют инсулин, макрофагный колониестимулирующий фактор, тромбоцитарный производный фактор роста. Например, мембранный рецептор инсулина, он является гликопротеином, который состоит из 2 α и 2 β субъединиц связанных дисульфидными связями. α субъединицы связывают инсулин, а β субъединицы обладают тирозинкиназной активностью. После присоединения гормона к α субъединицам, β субъединицы сначала фосфорилируют друг друга, а затем белок IRS-1 (insulin receptor substrate), который активирует функциональные ферменты (фосфопротеинфосфатаза). ФПФ в свою очередь дефосфорилирует и инактивирует инсулиновый рецептор.
б). Рецепторы, с фосфатазной активностью (тирозиновые протеинфосфотазы). Это каталитические рецепторы, дефосфорилирующие по тирозину белки-мишени (например, ФПФ).
в). Рецепторы с гуанилатциклазной активностью (гуанилатциклазы ГЦ). Это каталитические рецепторы, превращающие ГТФ в цГМФ, есть в сердце, легких, почках, надпочечниках, эндотелии кишечника, сетчатке и т.д.
Эти рецепторы находятся на мембране и в цитоплазме:
Мембранная ГЦ – гликопротеин (180кДа), имеет 3 домена: внеклеточный рецепорный, трансмембранный и внутриклеточный каталитический. Активируется предсердным натрийуретическим фактором (АНФ), термостабильным токсином грамотрицательных бактерий, эндотелийпроизводным фактором, ацетилхолином+Са2+, серотонином, гистамином и т.д. Существует 3 вида.
Цитоплазматическая ГЦ состоит из α и β субъединиц и содержит гем, активируется оксидом азота NO (а также Н2О2, О2, жирными кислотами и продуктами ПОЛ).
3). Рецепторы, сопряженные с G-белками (GPCR от англ. G – protein coupled receptor), по строению их еще называют серпантинными. Это мономерные интегральные мембранные белки, полипептидная цепь которых 7 раз пронизывает клеточную мембрану. Внеклеточный домен GPCR обеспечивает взаимодействие с гормоном, а внутриклеточный – контакт с G-белками. В настоящее время открыто более 200 видов GPCR. К GPCR относятся: α и β рецепторы. К GPCR присоединяются: адреналин (рецепторы α1 и α2, β1 и β2), ацетилхолин (рецепторы М1, М2, М3, М4), серотонин (1А, 1В, 1С, 2), дофамин (Д1 и Д2), АКТГ, ТТГ, ФСГ, ЛГ, хорионический гонадотропин, простагландины, гастрин, холецистокинин, нейропептид Y, нейромедин К, вазопрессин, ангиотензин, вещество К, вещество Р, или нейрокинин 1, 2 и 3 типа, тромбин, интерлейкин-8, глюкагон, кальцитонин, секретин, соматолиберин, ВИП, гипофизарный аденилатциклазактивирующий пептид, глютамат (MG1 – MG7), аденин.
GPCR через G-белки активируют аденилатциклазную и инозитолтрифосфатную системы. GPCR инактивируются:
путем ухода сигнальной молекулы из активного центра рецептора;
путем фосфорилирования рецепторов протеинкиназами и последующего присоединения специального белка (например, β-аррестин);
GPCR участвуют почти во всех жизненно важных процессах, протекающих в организме: они поддерживают работу сердца, органов пищеварения и дыхания, мозговую деятельность. Примерно половина современных лекарств взаимодействуют с GPCR.
4). Ядерные и цитоплазматические рецепторы. Находятся в цитоплазме или ядре, при взаимодействии с гормоном, образуют комплекс, который присоединяется к регуляторной нуклеотидной последовательности в ДНК, изменяет доступность промотора для РНК-полимеразы, соответственно меняет скорость транскрипции мРНК и трансляции новых ферментов. Ядерные и цитоплазматические рецепторы содержат ДНК-связывающий домен, характеризующийся наличием двух структур «цинковых пальцев». Особенностью цитоплазматических рецепторов является связь с белком-шапероном, который стабилизирует их структуру. Разные клетки организма, в зависимости от функций, имеют определенный набор рецепторов. На мембране одной клетки может быть более десятка разных типов рецепторов.
Регуляторные белки
G-белки – универсальные посредники, передающие сигнал от рецепторов к ферментам клеточных мембран.
В настоящее время известно более 50 G-белков:
Gs-белок активирует аденилатциклазу. Масса 80000-90000Да.
Gi-белок ингибирует аденилатциклазу. Масса 80000-90000Да. Через рецептор, активируется соматостатином.
Gq-белок активирует фосфолипазу С.
G-белки влияют на активность фосфодиэстеразы, фосфолипазы А2, некоторые типы Са2+- и K+-каналов.
G-белки также обеспечивают передачу сигнала в сенсорных клетках (фоторецепторных, обонятельных и вкусовых): Свет → родопсин → Gt → ФДЭцГМФ → (цГМФ→ГМФ) G-белки олигомеры, состоят из 3 субъединиц α, β, γ. β-субъединицы (35000 Да) у Gs- и Gi-белков одинаковы. α- субъединицы (41000 Да у Gi, 45000 Да у Gs) кодируются разными генами и обеспечивают специфический ответ (“+” или “-”).
Гормон (Г), взаимодействуя с рецептором (R), изменяет его конформацию.
Гормон-рецепторный комплекс, взаимодействуя с G-белком, уменьшает у α-субъединицы (α) сродство к ГДФ и увеличивает сродство к ГТФ.
Присоединение ГТФ к α-субъединице (в присутствии Mg2+) вызывает в G-белке изменение конформации и диссоциацию его на субъединицы: α-субъединицу (α-ГТФ) и димер βγ. α-ГТФ имеет высокое сродство к аденилатциклазе (Ац), его присоединение приводит к активации последней.
α-субъединица катализирует распад ГТФ до ГДФ + Фн. α-ГДФ имеет низкое сродство к Ац и высокое к димеру βγ. Отделение α-ГДФ от Ац инактивирует последнюю.
STAT белки. Вторичные посредники (мессенджеры)
Мессенджеры – низкомолекулярные вещества, переносящие сигналы гормонов внутри клетки. Они обладают высокой скоростью перемещения, расщепления или удаления (Са2+, цАМФ, цГМФ, ДАГ, ИТФ). Нарушения обмена мессенджеров приводят к тяжелым последствиям. Например, форболовые эфиры, которые являются аналогами ДАГ, но в отличие от которого в организме не расщепляются, способствуют развитию злокачественных опухолей.
цАМФ открыта Сазерлендом в 50 годах прошлого века. За это открытие он получил Нобелевскую премию. цАМФ участвует в мобилизации энергетических запасов (распад углеводов в печени или триглицеридов в жировых клетках), в задержке воды почками, в нормализации кальциевого обмена, в увеличении силы и частоты сердечных сокращений, в образовании стероидных гормонов, в расслаблении гладких мышц и так далее.
цГМФ активирует ПК G, ФДЭ, Са2+-АТФазы, закрывает Са2+-каналы и снижает уровень Са2+ в цитоплазме.
Ферменты
Ферменты каскадных систем катализируют:
образование вторичных посредников гормонального сигнала;
активацию и ингибирование других ферментов;
превращение субстратов в продукты;
Аденилатциклаза (АЦ)
Гликопротеин с массой от 120 до 150 кДа, имеет 8 изоформ, ключевой фермент аденилатциклазной системы, с Mg2+ катализирует образование вторичного посредника цАМФ из АТФ. АЦ содержит 2 –SH группы, одна для взаимодействия с G-белком, другая для катализа. АЦ содержит несколько аллостерических центров: для Mg2+, Mn2+, Ca2+, аденозина и форсколина. Есть во всех клетках, располагается на внутренней стороне клеточной мембраны. Активность АЦ контролируется: 1) внеклеточными регуляторами – гормонами, эйкозаноидами, биогенными аминами через G-белки; 2) внутриклеточным регулятором Са2+ (4 Са2+-зависимые изоформы АЦ активируются Са2+).
Протеинкиназа А
(ПК А) ПК А есть во всех клетках, катализируют реакцию фосфорилирования ОН- групп серина и треонина регуляторных белков и ферментов, участвует в аденилатциклазной системе, стимулируется цАМФ. ПК А состоит из 4 субъединиц: 2 регуляторных R (масса 38000 Да) и 2 каталитических С (масса 49000 Да). Регуляторные субъединицы имеют по 2 участка связывания цАМФ. Тетрамер не обладает каталитической активностью. Присоединение 4 цАМФ к 2 субъединицам R приводит к изменению их конформации и диссоциации тетрамера. При этом высвобождаются 2 активные каталитические субъединицы С, которые катализируют реакцию фосфорилирования регуляторных белков и ферментов, что изменяет их активность.
Протеинкиназа С (ПК С)
ПК С участвует в инозитолтрифосфатной системе, стимулируется Са2+, ДАГ и фосфатидилсерином. Имеет регуляторный и каталитический домен. ПК С катализирует реакцию фосфорилирования белков-ферментов.
Протеинкиназа G (ПК G)
Есть только в легких, мозжечке, гладких мышцах и тромбоцитах, участвует в гуанилатциклазной системе. ПК G содержит 2 субъединицы, стимулируется цГМФ, катализирует реакцию фосфорилирования белков-ферментов.
Фосфолипаза С (ФЛ С)
Гидролизует фосфоэфирную связь в фосфатидилинозитолах с образованием ДАГ и ИФ3, имеет 10 изоформ. ФЛ С регулируется через G-белки и активируется Са2+.
Фосфодиэстеразы (ФДЭ)
ФДЭ превращает цАМФ и цГМФ в АМФ и ГМФ, инактивируя аденилатциклазную и гуанилатциклазную систему. ФДЭ активируется Са2+, 4Са2+-кальмодулином, цГМФ.
NO-синтаза – это сложный фермент, представляющий собой димер, к каждой из субъединиц которого присоединено несколько кофакторов. NO-синтаза имеет изоформы.
Синтезировать и выделять NO способно большинство клеток организма человека и животных, однако наиболее изучены три клеточные популяции: эндотелий кровеносных сосудов, нейроны и макрофаги. По типу синтезирующей ткани NO-синтаза имеет 3 основные изоформы: нейрональную, макрофагальную и эндотелиальную (обозначаются соответственно как NO-синтаза I, II и III).
Нейрональная и эндотелиальная изоформы NO-синтазы постоянно присутствуют в клетках в небольших количествах, и синтезируют NO в физиологических концентрациях. Их активирует комплекс кальмодулин-4Са2+. NO-синтаза II в макрофагах в норме отсутствует. При воздействии на макрофаги липополисахаридов микробного происхождения или цитокинов они синтезируют огромное количество NO-синтазы II (в 100-1000 раз больше чем NO-синтазы I и III), которая производит NO в токсических концентрациях. Глюкокортикоиды (гидрокортизон, кортизол), известные своей противовоспалительной активностью, ингибируют экспрессию NO-синтазы в клетках.
Действие NO
NO – низкомолекулярный газ, легко проникает через клеточные мембраны и компоненты межклеточного вещества, обладает высокой реакционной способностью, время его полураспада в среднем не более 5 с, расстояние возможной диффузии небольшое, в среднем 30 мкм.
В физиологических концентрациях NO оказывает мощное сосудорасширяющее действие:
эндотелий постоянно продуцирует небольшие количества NO.
При различных воздействиях – механических (например, при усилении тока или пульсации крови), химических (липополисахариды бактерий, цитокины лимфоцитов и кровяных пластинок и т.д.) – синтез NO в эндотелиальных клетках значительно повышается.
NO из эндотелия диффундирует к соседним гладкомышечным клеткам стенки сосуда, активирует в них гуанилатциклазу, которая синтезирует через 5сек цГМФ.
цГМФ приводит к снижению уровня ионов кальция в цитозоле клеток и ослаблению связи между миозином и актином, что и позволяет клеткам через 10 с расслабляться.
На этом принципе действует препарат нитроглицерин. При расщеплении нитроглицерина образуется NO, приводящий к расширению сосудов сердца и снимающий в результате этого чувство боли.
NO регулирует просвет мозговых сосудов. Активация нейронов какой-либо области мозга приводит к возбуждению нейронов, содержащих NO-синтазу, и/или астроцитов, в которых также может индуцироваться синтез NO, и выделяющийся из клеток газ приводит к локальному расширению сосудов в области возбуждения.
NO участвует в развитии септического шока, когда большое количество микроорганизмов, циркулирующих в крови, резко активируют синтез NO в эндотелии, что приводит к длительному и сильному расширению мелких кровеносных сосудов и как следствие – значительному снижению артериального давления, с трудом поддающемуся терапевтическому воздействию.
В физиологических концентрациях NO улучшает реологические свойства крови:
NO, образующийся в эндотелии, препятствует прилипанию лейкоцитов и кровяных пластинок к эндотелию и также снижает агрегацию последних.
NO может выступать в роли антиростового фактора, препятствующего пролиферации гладкомышечных клеток стенки сосудов, важного звена в патогенезе атеросклероза.
В больших концентрациях NO оказывает на клетки (бактериальные, раковые и т.д) цитостатическое и цитолитическое действие следующим образом:
при взаимодействии NO с радикальным супероксид анионом образуется пероксинитрит (ONOO-), который является сильным токсичным окислителем;
NO прочно связывается с геминовой группой железосодержащих ферментов и ингибирует их (ингибирование митохондриальных ферментов окислительного фосфорилирования блокирует синтез АТФ, ингибирование ферментов репликации ДНК способствуют накоплению в ДНК повреждений).
NO и пероксинитрит могут непосредственно повреждать ДНК, это приводит к активации защитных механизмов, в частности стимуляции фермента поли(АДФ-рибоза) синтетазы, что еще больше снижает уровень АТФ и может приводить к клеточной гибели (через апоптоз).
Трансмембранная передача информации с участием аденилатциклазной системы
Последовательность событий, приводящих к каталитической активации ферментов
1). 1 Гормон (Г) присоединяется к Rs-рецептору с образованием гормон-рецепторного комплекса, который через несколько Gs-белков активирует несколько аденилатциклаз (комплекс гормон-Ri-рецептор через Gi-белки ингибирует аденилатциклазы);
2). 1 Аденилатциклаза превращает тысячи АТФ в тысячи цАМФ;
3). 4 цАМФ обратимо присоединяясь к 2 рецепторным субъединицам R ПК А, вызывают диссоциацию протомеров ПК А с освобождением 2 активных каталитических субъединиц С и двух 2цАМФ-R;
4). 1 Субъединица С ПК А фосфорилирует сотни ферментов или регуляторных белков, что способствует их активации или ингибированию. Таким образом, сигнал 1 гормона в аденилатциклазной системе суммарно усиливается в 106-107 раз;
5). Миллионы активных ферментов превращают молекулы субстрата в продукты. Инактивация аденилатциклазной системы осуществляется через ФДЭ и ФПФ. ФДЭ разрушает цАМФ до АМФ, это приводит к самосборке субъединиц ПК А и ее инактивации. ФПФ инактивирует (активирует) фермент в реакции гидролитического дефосфорилирования.
Посредством стимуляции аденилатциклазной системы оказывают гормональное действие кортиколиберин, кальцитонин, соматолиберин, ВИП, глюкагон, вазопрессин (через V2 рецепторы), ЛГ, ФСГ, ТТГ, хорионический гонадотропин, АКТГ, паратгормон, простагландины типа Е, D и I, адренергические катехоламины (через α2, β1и β 2 рецепторы).
Аденилатциклазная система активируется:
Угнетают аденилатциклазную систему гормоны: (через Gs-белки соматостатин), ангиотензин II, ацетилхолин (мускариновый эффект), дофамин, опиоиды и a2-адренергические катехоламины.
Трансмембранная передача информации с участием инозитолтрифосфатной системы
Последовательность событий, приводящих к каталитической активации ферментов
1). Гормон (Г) присоединяется к R-рецептору с образованием гормон-рецепторного комплекса, который через G-белок активирует фосфолипазу С;
2). Фосфолипаза С расщепляет фосфатидилинозитол-4,5-дифосфат (ФИФ2) клеточной мембраны на инозитолтрифосфат (ИТФ) и диацилглицерин (ДАГ);
3). ИФ3 присоединяясь к рецептору на мембране ЭПР, открывает кальциевые каналы, выпуская Са2+ из ЭПР в цитоплазму;
4). Са2+, ДАГ и фосфотидилсерин активируют ПК С, которая превращает субстрат в продукт.
5). 4 Са2+ присоединяются к кальмодулину, способствуя присоединению кальмодулина к ферменту и образованию активного комплекса 4Са2+-кальмодулин-фермент, который превращает субстрат в продукт. Инактивация инозитолтрифосфатной системы осуществляется кальциевыми насосами, которые откачивают Са2+ из цитоплазмы.
Посредством стимуляции инозитолтрифосфатной системы оказывают гормональное действие гонадолиберин, тиролиберин, дофамин, тромбоксаны А2, эндоперекиси, лейкотриены, агниотензин II, эндотелин, паратгормон, нейропептид Y, адренергические катехоламины (через α1 рецепторы), ацетилхолин, брадикинин, вазопрессин (через V1 рецепторы).
Инозитолтрифосфатная система активируется:
Трансмембранная передача информации с участием гуанилатциклазной системы
Последовательность событий, приводящих к каталитической активации ферментов
1). Гуанилатциклазная система функционирует в легких, почках, кишечнике, сердце, надпочечниках, эндотелии кишечника, сетчатке и др. Она участвует в регуляции водно-солевого обмена и тонуса сосудов, вызывает релаксацию и т.д.
Увеличение давления крови → предсердие → АНФ → почки → цГМФ → усиление секреции Na+ и H2O → усиление выведения из организма Na+ и H2O → снижение давления крови
Увеличение давления крови → предсердие → АНФ → ГМК сосудов → цГМФ → расслабление гладких мышц сосудов → снижение давления крови
Грамотрицательные бактерии → термостабильный токсин → эндотелий кишечника → цГМФ → торможение всасывания воды в кишечнике → диарея
Через образование оксида азота реализует свое действие ряд очень эффективных средств для лечения стенокардии (нитраты) и корректоров эректильной дисфункции (например, известный препарат Виагра).
Трансмембранная передача информации с участием цитоплазматических и ядерных рецепторов
Через цитоплазматические и ядерные рецепторы действуют кортикоиды, половые и тиреоидные гормоны.
Последовательность событий, приводящих к активации транскрипции и биосинтезу ферментов:
1). Освобождаясь от белка, гормон самостоятельно проходит клеточную мембрану и в цитоплазме присоединяется к цитоплазматическому рецептору с образованием гормон-рецепторного комплекса;
2). Гормон-рецепторный комплекс мигрирует в ядро, где присоединяется к регуляторной нуклеотидной последовательности в ДНК – энхансером или сайленсером.
3). При взаимодействии с энхансером увеличивается, а с сайленсером уменьшается доступность промотора для РНК-полимеразы, соответственно меняется скорость транскрипции мРНК и трансляции новых ферментов;
4). Ферменты превращают субстраты в продукты.
Иногда, гормон самостоятельно проникает в ядро, где соединяется с ядерным рецептором, образуя гормон-рецепторный комплекс. Этот комплекс также присоединяется к ДНК, запускает или блокирует процесс транскрипции мРНК и трансляции новых ферментов.
Изменение количества ферментов и интенсивности метаболизма под действием стероидных и териоидных гормонов происходит в течение нескольких часов.
Механизм действия ферментов.
Проблема строения и механизма действия ферментов насчитывает более 100 лет. Здесь будут рассмотрены лишь основные, этапные моменты ее развития. Белковая природа ферментов была окончательно установлена лишь в 30-х годах 20 века. Поэтому все представления о механизме действия ферментов до этого времени основывались преимущественно на исследованиях гомогенного химического катализа. Повышение скорости химических реакций под действием ферментов объяснялось активацией субстрата в образующемся фермент-субстратном комплексе.
Для объяснения высокой специфичности ферментов по отношению к определенным субстратам Э. Фишер в 1894г выдвинул гипотезу о строгом соответствии геометрической формы субстрата и активного центра фермента. Согласно предложенной им модели субстрат и фермент должны подходить друг к другу как ключ к замку:
Большой вклад в изучение механизма действия ферментов внесли Л. Михаэлис и М. Ментен (1913г), которые изучая кинетику ферментативных реакций пришли к выводу о том, что они включают несколько основных этапов:
Обратимое взаимодействие фермента с субстратом с образованием нестойкого фермент-субстратного комплекса;
Преобразование субстрата в составе фермент-субстратного комплекса;
Образование продуктов реакций, не обладающих сродством к ферменту, что способствует их освобождению: E + S ↔ E-S → E-P
Дальнейшим развитием теории Фишера явилось теория индуцированного (вынужденного) соответствия Д. Е. Кошланда (1959г), которая базировалась на данных исследований, включавших не только кинетический анализ, но и химическую модификацию радикалов аминокислот в молекуле фермента, ингибирование ферментов разными соединениями, изучение фермент-субстратных комплексов методами ренгено-структурного анализа, спектрографии и кристаллографии. Согласно этой теории полное соответствие центра и субстрата наступает лишь в процессе взаимодействия фермента с субстратом, который вызывает (индуцирует) необходимые конформационные изменения фермента.
Гипотеза «индуцированного соответствия» предполагает существование между ферментом и субстратом не только геометрического, но и электростатического соответствия, обусловленного спариванием противоположно заряженных групп субстрата и активного центра фермента, что является условием образования активного комплекса. В настоящее время считается установленным, что «индуцированное соответствие» фермента и субстрата достигается не только за счет конформационных изменений фермента, но и сопровождается геометрической и электронной перестройкой субстрата.
Определенный вклад в развитие представлений о ферментативном катализе внесла кислотно-основная теория катализа. Но дальнейшие исследования показали, что подход к рассмотрению действия ферментов с позиций обычного кислотно-основного катализа является упрощенным. В активном центре фермента, как правило, наличествует несколько реакционных центров, поэтому более правильно подходить к оценке их действия с позиции полифункционального катализа.
Все эти выводы включает в себя современная теория промежуточных соединений, согласно которой фермент при взаимодействии с субстратом образует комплекс, в котором реакционная способность исходных компонентов выше, чем в нативном состоянии. Через ряд промежуточных соединений происходит превращение субстрата в продукты реакции, при этом могут реализоваться разные механизмы катализа (кислотно-основной, электрофильный, нуклеофильный).
Кинетика ферментативных реакций.
Ферментативная кинетика изучает влияние различных факторов (концентрация S и E, рН, температура, давление, ингибиторы и активаторы) на скорость ферментативных реакций. Главной целью изучения кинетики ферментативных реакций является получение информации, позволяющей глубже понять механизм действия ферментов.
Кинетическая кривая позволяет определить начальную скорость реакции V0.
Кривая субстратного насыщения.
График зависимости V от концентрации субстрата при фиксированной концентрации фермента представляет собой гиперболу. Вначале скорость реакции прямо пропорциональна концентрации субстрата [S] (кинетика первого порядка). Однако при увеличении [S] скорость постепенно достигает максимального значения VMAX. Это означает, что все связывающие участки фермента заняты (насыщены). Скорость реакции на этом участке не зависит от концентрации субстрата (кинетика нулевого порядка). Такую кривую называют кривой субстратного насыщения.
Зависимость скорости реакции от концентрации фермента.
При постоянной концентрации субстрата существует прямо пропорциональная зависимость между скоростью реакции и концентрацией фермента [E] в реакционной смеси. Другими словами, для данной концентрации субстрата скорость реакции возрастает в 2 раза при двукратном увеличении концентрации фермента.
Зависимость скорости реакции от температуры.
Ферменты – вещества белковой природы, проявляют максимальную активность в ограниченном температурном режиме. При температурах не выше 40-50С скорость реакции увеличивается согласно теории химической кинетики. При более высоких температурах тепловая денатурация фермента приводит к полному прекращению ферментативной реакции. Термолабильность ферментов отличает ферменты от неорганических катализаторов.
Зависимость скорости реакции от рН.
Оптимум рН действия большинства ферментов лежит в пределах физиологических значений 6,0-8,0. Пепсин активен при рН 1,5-2,0, что соответствует кислотности желудочного сока. Аргиназа, специфичный фермент печени, активен при 10,0. Влияние рН среды на скорость ферментативной реакции связывают с состоянием и степенью ионизации ионогенных групп в молекуле фермента и субстрата. Этот фактор определяет конформацию белка, состояние активного центра и субстрата, формирование фермент-субстратного комплекса, собственно процесс катализа.
Математическое описание кривой субстратного насыщения, константа Михаэлиса.
Уравнение, описывающее кривую субстратного насыщения, было предложено Михаэлисом и Ментон и носит их имена (уравнение Михаэлиса-Ментен):
V = (VMAX*[S])/(Km+[S]), где Km – константа Михаэлиса. Легко рассчитать, что при V = VMAX/2 Km = [S], т.е. Km – это концентрация субстрата, при которой скорость реакции составляет ½ VMAX.
С целью упрощения определения величины VMAX и Km уравнение Михаэлиса-Ментен можно пересчитать. 1/V = (Km+[S])/(VMAX*[S]), 1/V = Km/(VMAX*[S]) + 1/VMAX,
1/V = Km/VMAX*1/[S] + 1/VMAX уравнение Лайнуивера-Берка. Уравнение, описывающее график Лайнуивера-Берка – это уравнение прямой линии (y = mx + c), где 1/VMAX – это отрезок, отсекаемый прямой на оси ординат; Km/VMAX – тангенс угла наклона прямой; пересечение прямой с осью абсцисс дает величину 1/Km. График Лайнуивера-Бэрка позволяет определить Km по относительно небольшому числу точек. Этот график также используют при оценке действия ингибиторов, о чем будет сказано ниже. Значение Km изменяются в широких пределах: от 10-6 моль/л для очень активных ферментов, до 10-2 – для малоактивных ферментов.
Оценки Km имеют практическую ценность. При концентрациях субстрата в 100 раз превышающих Km, фермент будет работать практически с максимальной скоростью, поэтому максимальная скорость VMAX будет отражать количество присутствующего активного фермента. Это обстоятельство используют для оценки содержания фермента в препарате. Кроме того, Km является характеристикой фермента, что используется для диагностики энзимопатий.
Ингибирование активности ферментов.
Чрезвычайно характеристикой и важной особенностью ферментов является их инактивация под влиянием определенных ингибиторов.
Ингибиторы – это вещества, вызывающие частичное или полное торможение реакций, катализируемых ферментами. Ингибирование ферментативной активности может быть необратимым или обратимым, конкурентным или неконкрентным.
Необратимое ингибирование – это стойкая инактивация фермента, возникающая в результате ковалентного связывания молекулы ингибитора в активном центре или в другом особом центре, изменяющим конформацию фермента. Диссоциация столь устойчивых комплексов с регенерацией свободного фермента практически исключена. Для преодоления последствий такого ингибирования организм должен синтезировать новые молекулы фермента.
Обратимое ингибирование – характеризуется равновесным комплексообразованием ингибитора с ферментом за счет нековалентных связей, вследствие чего такие комплексы способны к диссоциации с восстановлением активности фермента. Классификация ингибиторов на конкурентные и неконкурентные основана на том, ослабляется (конкурентное ингибирование) или не ослабляется (неконкурентное ингибирование) их ингибирующие действие при повышении концентрации субстрата.
Конкурентные ингибиторы – это, как правило, соединения, структура которых сходна со структурой субстрата. Это позволяет им связываться в том же активном центре, что и субстраты, препятствуя взаимодействию фермента с субстратом уже на стадии связывания. После связывания ингибитор может быть превращен в некий продукт или остается в активном центре, пока не произойдет диссоциация. Обратимое конкурентное ингибирование можно представить в виде схемы:
Степень ингибирования фермента определяется соотношением концентраций субстрата и фермента. Классическим примером подобного типа ингибирования является торможение активности сукцинатдегидрогеназы (СДГ) малатом, который вытесняет сукцинат из субстратного участка и препятствует его превращению в фумарат:
Ковалентное связывание ингибитора в активном центре приводит к инактивации фермента (необратимое ингибирование). Примером необратимого конкурентного ингибирования может служить инактивация триозофосфатизомеразы 3-хлорацетолфосфатом. Этот ингибитор является структурным аналогом субстрата – диоксиацетонфосфата и необратимо присоединяется к остатку глутаминовой кислоты в активном центре:
Некоторые ингибиторы действуют менее избирательно, взаимодействуя с определенной функциональной группой в составе активного центра разных ферментов. Так, связывание йодацетата или его амида с SH-группой аминокислоты цистеина, находящийся в активном центре фермента и принемающей участие в катализе, приводит к полной утрате активности фермента:
R-SH + JCH2COOH → HJ + R-S-CH2COOH
Поэтому эти ингибиторы инактивируют все ферменты, которые имеют SH-группы, участвующие в катализе. Необратимое ингибирование гидролаз при действии нервно-паралитических газов (зарин, зоман) обусловлено их ковалентным связыванием с остатком серина в активном центре.
Метод конкурентного ингибирования нашел широкое применение в медицинской практике. Сульфаниламидные препараты – антагонисты п-аминобензойной кислоты, могут служить примером метаболизируемых конкурентных ингибиторов. Они связываются с дигидроптератсинтетазой – бактериальным ферментом, осуществляющим превращение п-аминобензоата в фолиевую кислоту, необходимую для роста бактерий. Бактерия погибает в результате того, что связавшийся сульфаниламид превращается в другое соединение и фолиевая кислота не образуется.
Неконкурентные ингибиторы обычно связываются с молекулой фермента в участке, отличном от места связывания субстрата, и субстрат непосредственно не конкурирует с ингибитором. Поскольку ингибитор и субстрат связываются с разными центрами возможно образование как комплекса E-I, так и комплекса S-E-I. Комплекс S-E-I тоже распадается с образованием продукта, однако с меньшей скоростью, чем E-S, поэтому реакция будет замедляться, но не остановится. Таким образом, могут протекать следующие параллельные реакции:
Обратимое неконкурентное ингибирование встречается сравнительно редко. Неконкурентные ингибиторы называют аллостерическими в отличие от конкурентных (изостерических). Обратимое ингибирование может быть количественно изучено на основе уравнения Михаэлиса-Ментен.
При конкурентном ингибировании VMAX остается постоянной, а Km возрастает.
При неконкурентном ингибировании снижается VMAX при неизменном Km.
Если продукт реакции ингибирует фермент, катализирующий его образование, такой способ ингибирования называется ретроингибированием или ингибированием по принципу обратной связи. Например, глюкоза тормозит глюкозо-6-фосфатазу, которая катализирует гидролиз глюкозо-6-фосфата. Биологическое значение такого ингибирования – регуляция определенных метаболических путей (см. следующее занятие).
Медицинская энзимология
Энзимология – это раздел биохимии, изучающий ферменты и катализируемые ими реакции. Медицинская энзимология – это энзимология, которая изучает применение ферментов в медицине.
В медицинской энзимологии выделяют три основных направления:
энзимопатология;
энзимодиагностика;
энзимотерапия.
I. Энзимопатология
Энзимопатология – это наука, которая изучает энзимопатии. Энзимопатии – это группа заболеваний, которые вызваны различными дефектами ферментов.
Энзимопатий делятся на: наследственные (первичные) и приобретенные (вторичные).
1. Наследственные энзимопатии
Наследственные энзимопатии – это заболевания, вызванные наследственными нарушениями биосинтеза ферментов или их структуры и функции. В норме:
Полное или частичное нарушения биосинтеза ферментов вызывают дефекты генов регуляторных белков, которые контролируют синтез ферментов:
Нарушение структуры и функции ферментов вызывают дефекты генов этих ферментов:
У образовавшегося фермента наблюдаются структурные изменения, которые проявляются в изменении его каталитической активности (как правило, она исчезает), чувствительности к активаторам и ингибиторам, сродству к субстратам, оптимумам рН, температуры. В связи с этим изучением констант фермента является решающим в постановке диагноза врожденных энзимопатий.
Наследственные энзимопатии по типу нарушений метаболизма делят на:
1. нарушения обмена аминокислот: фенилкетонурия, альбинизм, алкаптонурия и др.;
2. нарушения углеводного обмена: галактоземия, наследственная непереносимость фруктозы, гликогенозы;
3. нарушения липидного обмена: липидозы;
4. нарушения обмена нуклеиновых оснований: подагры, синдрома Леш-Нихана и др.;
5. нарушение обмена в соединительной ткани: мукополисахаридозы, хондродистрофия и др.;
6. дефекты ферментов в ЖКТ: муковисцидоз, целиакия, непереносимость лактозы и др.
7. нарушения обмена стероидов и т.д.
В норме метаболический путь протекает следующим образом:
Из-за дефекта в метаболическом пути (цикле, шунте) одного из ферментов в организме происходит накопление промежуточных продуктов (часто токсичных в высоких концентрациях) и дефицит жизненно необходимых конечных продуктов, что приводит к клиническим проявлениям:
Пример: фенилпировиноградная олигофрения – наследственное заболевание, приводящее в раннем детстве к гибели ребенка или к развитию у него тяжелой умственной отсталости. Причиной заболевания является отсутствие в печени фермента фен-4-монооксигеназы, которая обеспечивает превращение незаменимой аминокислоты Фен в Тир:
Эта реакция необходима для катаболизма Фен, т.е. удаления его излишков. При отсутствии фен-4-монооксигеназы в организме происходит накопление Фен и превращение его в различные производные: фенилпировиноградную, фенилмолочную и фенилуксусную кислоты. Фен и его производные в высоких концентрациях токсичны, накапливаясь в тканях, они оказывают на них повреждающее действие. Самой чувствительной к Фен и его производным оказывается нервная ткань детей, она поражается в первую очередь.
Диагноз фенилкетонурия ставят на основании обнаружения Фен в крови или фенилпировиноградной кислоты на пеленках детей. Лечение в основном сводится к исключению из питания ребенка Фен. Для такого ребенка Тир оказывается незаменимой аминокислотой.
Другое тяжелое наследственное заболевание – галактеземия (непереносимость молочного сахара), связано с отсутствием синтеза в печени ферментов, катализирующих превращение галактозы в глюкозу. В результате в раннем возврате происходит накопление в тканях галактозы, приводящее к развитию катаракты, поражению печени, мозга, нередко вызывающее гибель ребенка. Лечение в данном случае сводиться к исключению из диеты молочного сахара.
2. Приобретенные энзимопатии
Приобретенные энзимопатии делятся на: алиментарные, токсические и вызванные различными патологическими состояниями организма.
А). Алиментарные энзимопатии – это заболевания, вызванные изменением количества и активности ферментов вследствие нарушения характера питания.
Алиментарные энзимопатии вызываются дефицитом или дисбалансом в пище:
витаминов (гипо-, авитаминозы);
макро- и микроэлементов;
аминокислот;
жирных кислот;
других БАВ
Например, алиментарная энзимопатия, вызванная недостаточностью витамина А, проявляется нарушением сумеречного зрения (куриная слепота), воспалением слизистых глаз, ЖКТ, кожи.
Б). Токсические энзимопатии – это заболевания, вызванные нарушением активности ферментов вследствие действия токсических веществ. Токсическим веществами являются многие ксенобиотики (соли тяжелых металлов, пестициды, гербециды и т.д.), а также некоторые метаболиты в высоких концентрациях (алкоголь).
Токсические вещества могут либо избирательно угнетать активность (через денатурацию или ингибирование) или синтез отдельных ферментов, либо угнетать весь биосинтез белка (и соответственно всех ферментов).
Цианиды и СО прочно связываются с геминовым Fe активного центра цитохромов, что угнетает их активность;
Фториды угнетают активность ферментов, содержащих в активном центре Mg2+;
Ингибиторы, содержащиеся в соевых продуктах, яйцах домашней птицы угнетают активность протеаз ЖКТ – трипсина, химотрипсина, эластазы;
Антивитамины, присутствующие в некоторых пищевых продуктах, или разрушают витамины или конкурентно замещают их в молекулах ферментов, что приводит к угнетению активности этих ферментов.
В). Энзимопатии, вызванные различными патологическими состояниями организма. Так как ферменты имеют оптимумы t, рН и давления, практически любое заболевание, вызывающее нарушения КОС, изменение температуры тела, концентрации активаторов и ингибиторов, меняет активность ферментов организма. Например, при ацидозе и повышении температуры возрастает активность катаболических (лизосом) и падает активность анаболических ферментов.
Ферменты в медицинской практике находят применение в качестве диагностических (энзимодиагностика) и терапевтических (энзимотерапия) средств.
II Энзимодиагностика
Энзимодиагностика (энзим[ы] + греч. diagnostikos способный распознавать) – методы диагностики болезней, патологических состояний и процессов, основанные на определении активности ферментов в биологических жидкостях.
Направления энзимодиагностики:
Определение активности органо-, органеллоспецифических ферментов и их изоферментов.
Определение активности ферментов и их констант (Km, t, pH).
Определение концентрации органических веществ с помощью ферментов.
1) Определение активности органо-, органеллоспецифических ферментов и их изоферментов. Определение в биологических жидкостях активности ферментов и их изоферментов позволяет установить локализацию патологического процесса, его стадию, выраженность, а также эффективность его лечения.
Ферменты плазмы крови по происхождению можно разделить на 3 группы:
1) секреторные ферменты – секретируются определенными органами в плазму крови, где и выполняют свою функцию. Например: ЛПЛ, ЛХАТ, ферменты свертывающей и противосвертывающей системы крови;
2) экскреторные ферменты – синтезируются в железах ЖКТ, выделяются в просвет ЖКТ, где обеспечивают процесс пищеварения. В кровь эти ферменты попадают при повреждении желез. Например, при панкреатите в крови обнаруживается панкреатическая липаза, амилаза, трипсин и т.д., при воспалении слюнных желез – амилаза слюны.
3) клеточные ферменты – функционируют только внутри клеток, в плазму крови они попадают во время повреждения (под действием медиаторов воспаления и т.д.) и гибели клеток (при некрозе). К ним относятся общие, органо- и органеллоспецифические ферменты.
За счет естественной гибели клеток клеточные ферменты имеют в плазме крови постоянно низкую активность. При поражении органа происходит значительный выход ферментов из его клеток и многократное увеличение активности этих ферментов в плазме крови.
Аминотрансферазы. Локализуются в митохондриях, обеспечивают взаимопревращения аминокислот и кетокислот: АК1 + КК2 ↔ КК1 + АК2. АСТ: асп+α-КГ↔ЩУК+глу.
АСТ много в миокарде, по убыванию меньше в печени, скелетной мускулатуре, ЦНС, почках, семенниках. Активность в сыворотке крови 6-25МЕ/л. АЛТ: ала+α-КГ↔ПВК+глу.
АЛТ много в печени, поджелудочной железе, миокарде, скелетной мускулатуре. Активность в сыворотке крови 6-26МЕ/л. Активность обеих трансаминаз в сыворотке крови возрастает в десятки раз при инфаркте миокарда (АСТ>АЛТ), при остром инфекционном гепатите (АЛТ>АСТ), а также при циррозе печени и мышечной дистрофии.
Информативными пробами являются креатинфосфокиназный и лактатдегидрогеназный тесты, относящиеся к некротическим ферментным методам. Их диагностическая ценность повысилась после внедрения в клиническую практику методов определения их изоферментов.
ЛДГ. Локализуется в цитозоле, обеспечивает взаимопревращения ПВК и лактата. ЛДГ: ПВК+ НАДН2 ↔ лактат + НАД+. ЛДГ1 и ЛДГ2 наиболее активны в сердечной мышце и почках, ЛДГ4 ЛДГ5 – в скелетных мышцах и печени. Активность в сыворотке крови 55-140МЕ/л. При инфаркте миокарда в сыворотке крови резко повышается активность ЛДГ1 и ЛДГ2, а при поражениях скелетной мускулатуры и печени (гепатит, отравления хлорорганическими соединениями) повышается активность ЛДГ4 ЛДГ5.
Креатинкиназа (КК). Локализуется в цитозоле, митохондриях, миофибриллах. КК: Креатин + АТФ ↔ креатинфосфат + АДФ. Изофермент КК-ВВ находиться преимущественно в головном мозге, МВ – в миокарде, ММ – в скелетных мышцах. Активность в сыворотке крови КК в норме до 90МЕ/л.
В сыворотке крови КК-ММ повышается при повреждении скелетных мышц (при прогрессирующей мышечной дистрофии в 50 раз), КК-МВ – при инфаркте миокарда. КК-ВВ не проходит гематоэнцефалический барьер и не имеет значения для диагностики патологий ЦНС.
В сыворотке крови повышена активность амилазы при остром панкреатите, кисте поджелудочной железы; γ-глутамилтранспептидазы – при остром инфекционном или токсическом гепатите, хроническом гепатите, циррозе печени; кислой фосфатазы – при карциноме простаты; щелочной фосфатазы – при заболеваниях костей, закупорке желчных протоков, при беременности и у детей. Определение активности ферментов с диагностической целью проводят также в моче, слюне, ликворе и биоптатах органов и тканей.
2) Определение активности ферментов и их констант (Km, t, pH). Это направление используют для диагностики наследственных патологий и выявления механизмов патогенеза ряда заболеваний.
3) Определение концентрации органических веществ с помощью ферментов. Использование ферментов в качестве реактивов позволяет определять концентрацию органических веществ с высокой точностью, так как ферменты обладают высокой чувствительностью и избирательностью к своим субстратам.
III Энзимотерапия
Энзимотерапия – применение ферментов животного, бактериального или растительного происхождения и регуляторов активности ферментов с лечебной целью. Внедрению ферментных препаратов в современную клиническую практику способствовало развитие технологий получения обогащенных ферментами препаратов и очищенных ферментов.
В энзимотерапии существует насколько направлений:
1. Лекарственные препараты на основе ферментов. В качестве лекарственных препаратов наиболее широко используются гидролитические ферменты.
1). Протеолитические ферменты применяются при нарушении пищеварения.
а). Экстракты слизистой оболочки желудка, основным действующим веществом которых является пепсин. Это препараты абомин и ацидинпепсин, их в основном используют для коррекции секреторной дисфункции желудка.
б). Панкреатические энзимы, представленные амилазой, липазой, трипсином и химотрипсином. Это препараты панкреатин, мезим-форте, панцитрат, креон, их используются для коррекции нарушений процесса пищеварения, а также для регуляции функций поджелудочной железы.
в). Комбинированные ферменты, содержащие панкреатин в комбинации с компонентами желчи, гемицеллюлозой. Это препараты дигестал, фестал, панзинорм-форте, энзистал. Их назначают при недостаточной внешнесекреторной функции поджелудочной железы в сочетании с патологией печени, желчевыводящей системы, при нарушении жевательной функции, малоподвижном образе жизни, кратковременных погрешностях в еде.
г). Растительные энзимы, представленные папаином, грибковой амилазой, протеазой, липазой и др. ферментами. Препараты пепфиз и ораза. Папаин и протеазы гидролизируют белки, грибковая амилаза – углеводы, липаза – жиры.
д). Дисахаридазы. Например, тилактаза – пищеварительный фермент представляющий собой лактазу, которая находится в щеточной кайме слизистой оболочки тощей кишки и проксимального отдела подвздошной кишки. Расщепляет лактозу на простые сахара.
2). Протеолитические препараты применяют местно, в виде аппликаций или орошений, при первичной обработке ран и ожогов. Гидролизуя белки некротизированных тканей, ферменты способствуют очищению раны, уменьшению воспаления и ускорению заживления. Например, коллагеназа вызывает деструкцию коллагена при этом жизнеспособные мышцы, грануляционная ткань и эпителий остаются интактными. При гнойных ранах коллагеназа способствует быстрому очищению от нежизнеспособных тканей и экссудата, более раннему появлению грануляционной ткани и эпителизации, предупреждает развитие грубых (типа келоидных) рубцов, способствует сохранению функции суставов.
3). Гиалуронидазы – (лидаза, ронидаза) – ферменты, специфическим субстратом которых является гиалуронованная кислота, основа межклеточного матрикса соединительной ткани. Показателями к их применению являются рубцы после ожогов и операций, гематомы, контрактуры суставов и т.д. Лечебный эффект проявляется размягчением рубцов, рассасыванием гематом, появлением подвижности в суставах.
4). Протеолитические ферменты стрептокиназу, урокиназу применяют для предотвращения и лечения тромбозов, эмболии, инфаркта миокарда, закупорки сосудов сетчатки глаза. Стрептокиназа и урокиназа способствует превращению плазминогена в плазмин, который лизирует тромб.
5). Нуклеазы (ДНК-аза, РНК-аза) используют при лечении некоторых вирусных заболеваний (герпес, аденовирусный конъюктивит, вирусный менингит, ОРВИ и др.). Фермент разрушает ДНК вируса, не повреждая вместе с тем ДНК клеток макроорганизма. α-ДНКаза в препарате пульмозим используется как муколитик, она расщепляет внеклеточную ДНК, содержащуюся в большом количестве в вязком бронхиальном секрете.
6). Бактериолитические ферменты. Препарат «лизоамидаза» содержит комплекс ферментов: мурамидазу, амидазу, пептидазу и высокомолекулярный полисахарид. Препарат обладает наибольшей бактерицидной активностью по отношению к грамположительным бактериям: стафилакоккам, стрептококкам, а также менингококкам, гонококкам. Особенность, препарат показывает высокую бактерицидную эффективность вне зависимости от устойчивости бактерий к действию антибиотиков.
Системная энзимотерапия – использование полиферментных препаратов широкого спектра действия. Полиферментные препараты представляют собой эффективное противовоспалительное, противоотечное, фибринолитическое, иммуномодулирующее и вторично анальгетическое средство широкого спектра показаний. Препараты вобэнзим, вобэмугос, флогэнзим применяются в области травматологии, хирургии, спортивной медицины, ревматологии, ангиологии и гинекологии. Препарат вобэнзим кроме панкреатина содержит папаин (из растения Carica Papaya), бромелаин (из ананаса обыкновенного) и рутозид (группа витамина Р). Наряду с выраженными ферментативными свойствами обладает противовоспалительным, противоотечным, фибринолитическим и вторичноанальгезирующим действием. Используется при панкреатите, язвенном колите, болезни Крона, травмах, аутоиммунных онкологических, урологических, гинекологических заболеваниях.
Флогэнзим представляет собой комбинацию ферментов (бромелаин и трипсин) и рутина. Бромелаин и трипсин способствуют быстрому расщеплению клеточных фрагментов и метаболических продуктов воспалительного процесса, рутин восстанавливает проницаемость стенок сосудов, что приводит к уменьшению отеков и гематом. Флогэнзим оказывает противовоспалительное, фибринолитическое, антиагрегантное и противоотечное действие.
Применение ферментов в терапии ограничивается их лабильностью, нежелательными антигенными свойствами, трудностями адресной доставки к пораженным органам и тканям. Этих недостатков лишены иммобилизированные ферменты – энзимные препараты, соединенные с полимерной матрицей, либо включенные в липосомы или тени эритроцитов. Например, прозефим (комплекс бактериальных протеаз, иммобилизированных на аминоцеллюлозе), применяемый при гнойно-некротических процессах, обладает пролонгированным лечебным эффектом. Он способен гидролизовать денатурированный белок непрерывно свыше 10 суток, сохраняя по истечении этого срока до 90% исходной активности.
Иммобилизированные ферменты применяют для экстракорпоральной перфузии крови типа «искусственная почка». Такое лечение полностью исключает нежелательные воздействия на организм чужеродного белка и может проводиться в течение длительного времени.
II). Лекарственные препараты на основе коферментов и витаминов. Коферментные препараты: кокарбоксилаза, рибофлавина мононуклеотид, флавинат, пиридоксальфосфат, кобамамид используют для усиления активности ряда ферментов.
III). Лекарственные препараты на основе ингибиторов и активаторов ферментов.
Препараты пантрипин, ингитрил, контрикал ингибируют протеолитических ферменты поджелудочной железы, которые активируются в протоках при воспалении, и применяются для лечения острых панкреатитов.
Антихолинэстеразные препараты, ингибиторы моноаминоксидазы применяются как психотропные средства. ингибиторы карбоангидразы используются в качестве диуретиков.
Аллопуринол используют при гиперурикемии для ингибирования фермента ксантиноксидазы.
Действие тетурама при лечении алкоголизма связано с угнетением им фермента ацетальдегидоксидазы.
КЛАССИФИКАЦИЯ ФЕРМЕНТОВ
Современная номенклатура ферментов допускает старые тривиальные названия (папаин, пепсин, трипсин, цитохромы) и названия по рациональной номенклатуре, согласно которой название фермента составляется из названия субстрата и окончания –аза (амилаза, липаза, протеаза). По научной номенклатуре название фермента составляют из химического названия субстрата, акцептора и названия той реакции, которая осуществляется ферментом, например, L-аланин:2-оксоглутарат аминотрансфераза.
Существует Международная классификация ферментов. Согласно ей все ферменты включены в «Каталог ферментов» под своим классификационным номером, состоящим из четырех цифр. Первая цифра указывает на принадлежность к одному из шести главных классов. Следующие две определяют подкласс и подподкласс, а последняя цифра – номер фермента в данном подподклассе. Например, лактатдегидрогеназа имеет номер КФ 1.1.1.27.
В основу классификации ферментов положен тип реакций, подвергающихся каталитическому воздействию. Классы ферментов:
2. Трансферазы – переносят ту или иную функциональную группу от одного субстрата на другой.
3. Гидролазы – также участвуют в переносе групп, однако акцептором всегда является молекула воды.
4. Лиазы (синтазы) – катализируют расщепление или образование химических соединений или образование химических связей, при этом образуются или исчезают двойные связи.
5. Изомеразы – перемещают группы в пределах одной молекулы.
6. Лигазы (синтетазы) – катализируют энергозависимые реакции присоединения и поэтому их действие сопряжено с гидролизом АТФ.
Оксидоредуктазы катализируют реакции, имеющие общий вид:
Характерной особенностью деятельности оксидоредуктаз в клетке является их способность образовывать системы окислительно-восстановительных ферментов, в которых осуществляется многоступенчатый перенос атомов водорода от первичного субстрата к конечному акцептору, обычно кислороду, с образованием воды.
Оксидоредуктазы, которые переносят протоны и электроны непосредственно на кислород, называются аэробными дегидрогеназами или оксидазами. Оксидоредуктазы, переносящие протоны и электроны от одного компонента окислительной цепи к другому без передачи на кислород, называются анаэробными дегидрогеназами или редуктазами.
Если фермент катализирует реакцию отнятия атомов водорода непосредственно от первичного субстрата, то его называют первичной дегидрогеназой.
Если фермент ускоряет снятие атомов водорода со вторичного субстрата, который сам ранее получил атомы водорода при помощи первичной дегидрогеназы, то его называют вторичной дегидрогеназой.
Другая особенность оксидоредуктаз состоит в том, что они ускоряют протекание химических процессов, связанных с высвобождением энергии.
Третья особенность оксидоредуктаз заключается в том, что имея несколько коферментов, они способны ускорять большое количество разных окислительно-восстановительных реакций. Один и тот же кофермент способен соединяться со многими апоферментами, образуя каждый раз оксидоредуктазу, специфичную по отношению определенному субстрату.
Самыми распространенными являются оксидоредуктазы, содержащие в качестве активной группы никотинамидадениндинуклеотид (НАД + ) – производное никотинамида, витамина РР.
окисленная форма НАД + восстановленная форма НАДН+Н
Субстратами НАД-зависимых дегидрогеназ являются спирты, альдегиды, амины, дикарбоновые и кетокислоты. Все НАД-зависимые дегидрогеназы являются анаэробными дегидрогеназами, то есть посылают атомы водорода на ближайший в окислительной цепи другой фермент.
Примером НАД-зависимой дегидрогеназы является алкогольдегидрогеназа из печени животных. Фермент состоит из двух субъединиц, каждая из которых несет молекулу НАД + и атом Zn. Он катализирует реакцию окисления спирта в альдегид:
Кроме НАД + оксидоредуктазы содержат в качестве кофермента НАДФ + , у которого водород ОН-группы 2-го углеродного атома рибозы аденозина замещен на остаток фосфорной кислоты.
Партнером НАД-зависимых дегидрогеназ выступают флавопротеиды, например флавинмононуклеотид ФМН, производное витамина В2. Окисленная форма ФМН окрашена, она способна принимать два протона и два электрона, переходя в лейкоформу:
окисленная окрашенная форма ФМН восстановленная бесцветная форма ФМНН2
Другим коферментом во флавопротеинах является ФАД:
Флавопротеины в большинстве случаев являются вторичными дегидрогеназами, однако некоторые флавопротеины, особенно с ФАД, могут непосредственно снимать атомы водорода с субстрата. Примером реакции с участием ФАД является окисление глюкозы глюкозооксидазой в глюконо-лактон. Глюкозооксидаза синтезируется плесневелыми грибами при рН 5–7. Фермент содержит две молекулы ФАД, высоко специфичен и стабилен. Применяют для получения D-глюконата кальция и удаления О2 из продуктов.
Коферментами оксидоредуктаз также являются хиноны – убихинон дыхательной цепи, пластохинон фотосинтезирующей системы хлоропластов. Они передают атомы водорода по окислительно-восстановительным цепям:
Несколько оксидоредуктаз образуют цитохромную систему, имеющую в качестве простетических групп железопорфирины (геминовое железо). Они отличаются белковой частью. Обозначаются буквами a, b, c и d с индексами. Цитохромная система способна принимать электроны, снятые с восста-новленного убихинона. Она передает электроны далее по цепи цитохромов, в конце концов соединяя их с кислородом.
Трансферазы – один из наиболее обширных классов, специализи-руются на переносе функциональных групп с одного субстрата к другому. Делятся на подклассы в зависимости от характера переносимых групп.
Фосфотрансферазы – ускоряют реакцию переноса остатка фосфорной кислоты. Реакцией фосфорилирования обеспечивается превращение ряда органических соединений в фосфорные эфиры, обладающие повышенной химической активностью и более легко вступающие в последующие реакции. Перенос фосфатных групп идет на спиртовые, карбоксильные, азотсодержащие, фосфорсодержащие и другие группы. Донором фосфатных остатков является в большинстве случаев АТФ. Фосфотрансферазы, переносящие остаток фосфата с АТФ на белки, называются протеинкиназами. Они переносят фосфат на радикалы серина, треонина, тирозина, лизина и гистидина, в результате чего резко изменяется биологисческая активность ряда белков. Фосфотрансферазой является, например, гексокиназа – фермент, ускоряющий перенос остатка фосфорной кислоты от молекулы АТФ к глюкозе с образованием глюкозо-6-фосфата. Гексокиназа состоит из 4 субъединиц, каждая отдельная молекула лишена фосфотрансферазной активности. Обнаружено 5 изозимов гексокиназы:
глюкоза АТФ
Ацилтрансферазы – ускоряют перенос ацилов (остатков карбоновых кислот) на аминокислоты, амины, спирты и другие соединения. Универсальным источником ацильных групп во всех этих реакциях является ацилкоэнзим А (ацил-СоА), чаще всего переносу подвергается ацетил. Одним из примеров реакции трансацилирования является синтез ацетилхолина холинацетилтрансферазой:
ацетил-СоА холин ацетилхолин СоА
Аминотрансферазы – ускоряют реакцию трансаминирования аминокислот с кетокислотами и очень важны для обеспечения биосинтеза аминокислот. Простетической группой аминотрансфераз является пиридоксальфосфат, производное витамина В6 (пиридоксина). Механизм реакции трансаминирования хорошо изучен. Ниже приведено суммарное уравнение и механизм реакции трансаминирования аланина с помощью фермента аланинаминотрансферазы (АЛТ).
Аланин + α-кетоглутарат ↔ пируват + глутамат.
аланин пиридоксальфосфат основание Шиффа
α-кетоглутарат пиридоксаминфосфат основание Шиффа
Важное значение среди трансфераз имеют ферменты, ускоряющие перенос одноуглеродных фрагментов, содержащие в качестве коферментов фолиевую кислоту и цианокобаламин.
Гликозилтрансферазы – ускоряют реакции переноса гликозильных остатков из молекул фосфорных эфиров или других соединений к молекулам моносахаридов, полисахаридов или других веществ. Примером может служить фермент УДФ-глюкоза:гликоген-4-α-глюкозилтрансфераза, который осуществляет наращивание молекул гликогена на один остаток глюкозы:
УДФ-глюкоза нередуцирующий конец
увеличенный на один остаток глюкозы УДФ
нередуцирующий конец молекулы гликогена
Гидролазы– самая распространенная группа ферментов, не содержат коферментов, то есть являются по химической структуре простыми белками. В зависимости от характера гидролизуемого субстрата гидролазы делятся на ряд подклассов.
Эстеразы – катализируют реакции гидролиза сложных эфиров спиртов с органическими и неорганическими кислотами. Примером эстераз является липаза, которая ускоряет гидролиз сложноэфирных связей в молекулах жиров:
Другим примером являются фосфатазы, катализирующие гидролиз фосфорных эфиров, например глюкозо-1-фосфатаза:
Гликозидазы – ускоряют реакцию гидролиза гликозидов. Обладают ярко выраженной пространственной специфичностью (α- и β-гликозидазы). Примером может служить сахараза, ускоряющая гидролиз сахарозы на глюкозу и фруктозу:
сахароза глюкоза фруктоза
Пептидазы – катализируют расщепление пептидных связей в белках и пептидах. Эндопептидазы катализируют гидролиз пептидных связей внутри молекулы, экзопептидазы – отщепление от полипептидной цепи концевых аминокислот (аминопептидазы и карбоксипептидазы). Существуют также дипептидазы, завершающие гидролиз белка. В зависимости от меха-низма действия эндопептидазы делятся на четыре подподкласса:
а) сериновые протеиназы, несут в активном центре остатки серина
и гистидина (трипсин, химотрипсин, субтилизин);
б) тиоловые тротеиназы, содержащие в активном центре цистеин (папаин,
фицин, бломелаин, катепсин В);
в) кислые протеиназы, имеющие оптимум рН ниже 5 и содержащие радикалы
дикарбоновых аминокислот в активном центре (пепсин, катепсин D);
г) металлопротеиназы, каталитическое действие которых зависит от ионов
металлов Са 2+ и Zn 2+ в активном центре (коллагеназа, термолизин).
Амидазы – ускоряют гидролиз амидов кислот. Из них важную роль играют уреаза, аспарагиназа и глутаминаза.
Уреаза была первым ферментом, полученным в кристаллическом виде. Это простой фермент, молекула его глобулярна и состоит из 8 одинаковых субъединиц. Уреаза обладает абсолютной специфичностью и ускоряет гидролиз мочевина до NH3 и СО2.
Аспарагиназа и глутаминаза ускоряют гидролиз амидов аспартата и глутамата, например:
К гидролазам, действующим на C-N связи, относится аргиназа. Она обладает абсолютной специфичностью и гидролизует аминокислоту аргинин на орнитин и мочевину:
аргинин мочевина орнитин
Лиазы – ускоряют негидролитические реакции распада органических соединений по связям C-C, C-N, C-O, C-S. При этом замыкаются двойные связи и выделяются такие простейшие продукты, как CO2, H2O, NH3. Некоторые из этих реакций являются обратимыми, и соответствующие ферменты в подходящих условиях катализируют реакции не только распада, но и синтеза.
Одной из важнейших групп этого класса являются углерод-углерод-лиазы. К ним относятся декарбоксилазы кето- и α-аминокислот и альдегид-лиазы. Представителем альдегид-лиаз является альдолаза, катализирующая обратимую реакцию расщепления фруктозо-1,6-дифосфата до фосфотриоз:
В качестве коферментов декарбоксилазы кетокислот используют производное витамина В1 – тиаминпирофосфат, а декарбоксилазы α-амино-кислот – пиридоксальфосфат. Например, пируватдекарбоксилаза и лизин-декарбоксилаза катализируют следующие две реакции соответсвенно:
Другой важной группой лиаз являются углерод-кислород-лиазы, ускоряющие реакции гидратирования и дегидратирования при метаболизме углеводов и жирных кислот. Например, фумарат-гидратаза катализирует присоединение воды по двойной связи фумарата с образованием малата:
Примером углерод-азот-лиаз может служить аспартат-аммиак-лиаза, ускоряющая реакцию прямого дезаминирования аспартата, фермент характерен для бактерий и ряда растений:
Некоторые лиазы ускоряют реакции не только распада, но и синтеза. Например, из дрожжей выделен фермент L-серин-гидро-лиаза, отщепляющий от серина воду и присоединяющий сероводород, в результате чего синтезируется аминокислота цистеин. Подобные лиазы также называют синтазами. Следует отличать их от ферментов класса лигаз, которые ускоряют реакции только синтеза и именуются в связи с этим синтетазами. Серин-гидро-лиазу можно также назвать цистеинсинтазой:
Изомеразы – немногочисленный класс ферментов, осуществляют внутримолекулярный перенос водорода, фосфатных и ацильных групп, перемещение двойных связей, изменение пространственного расположения атомных группировок.
Триозофосфатизомераза ускоряет перенос атомов водорода в процессе взаимопревращений глицеральдегид-3-фосфата и дигидроксиацетонфосфата:
Фосфоглицератфосфомутаза обеспечивает достаточную скорость превращения 2-фосфоглицерата в 3-фосфоглицерат и обратно:
Мутаротаза является стереоизомеразой, она ускоряет реакцию превращения α-D-глюкопиранозы в β-D-глюкопиранозу. К стереоизомеразам относятся также цис-транс-изомеразы, например ретинол-изомераза, переводящая транс-ретинол в цис-ретинол.
Изопентенилпирофосфат-изомераза катализирует реакцию перестройки изопентенилпирофосфата в диметилаллилпирофосфат, что связано с переме-щением двойной связи из 3-го во 2-е положение:
Изопентенилпирофосфат-изомераза содержит свободные сульфгидрильные группы цистеина, благодаря которым обеспечивается приведенная выше реакция, имеющая огромное значение для синтеза изопреноидов.
Лигазы – ферменты, катализирующие синтез органических веществ в клетке. Главная их особенность – сопряженность синтеза с распадом АТФ. При отрыве от ее молекулы в присутствии лигаз одного или двух концевых остатков фосфорной кислоты выделяется большое количество энергии, используемой для активирования реагирующих веществ. Далее лигазы ускоряют соединение этих активированных субстратов.
К группе лигаз, образующих С-С-связи, относятся карбоксилазы. Например, пируваткарбоксилаза ускоряет реакцию образования оксалоацетата из пирувата и СО2:
В качестве кофермента карбоксилазы используют активированную форму витамина Н (биотина) – карбоксибиотин:
Другим примером работы лигаз может служить ускорение реакции образования C-S-связей, осуществляемое ацил-коэнзим А-синтетазами. Ацетил-коэнзим А-синтетаза катализирует образование ацетил-коэнзима А из уксусной кислоты и коэнзима А:
ацетат коэнзим А ацетил-СоА
Ацетил-коэнзим А служит коферментом в реакциях трансацилирова-ния, поэтому действие ферментов этих двух групп – лигаз и ацилтрансфераз – тесно связано друг с другом.
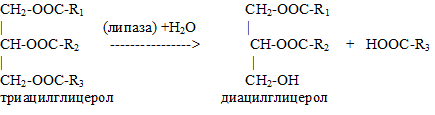
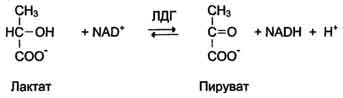
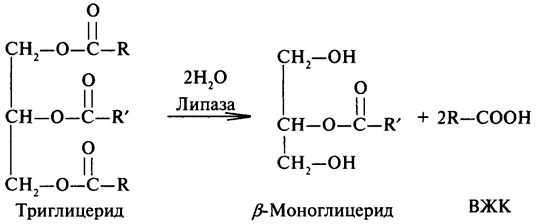
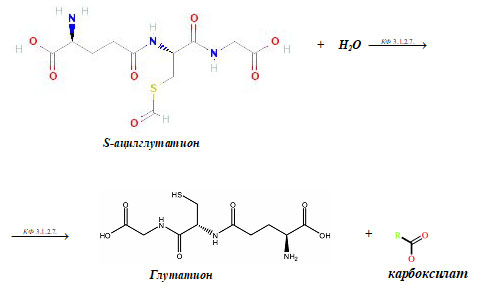




































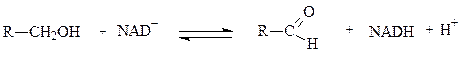
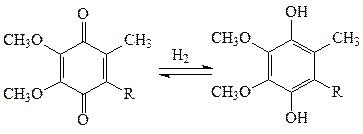
 глюкоза АТФ
глюкоза АТФ