Нуклеиновые кислоты способны гидролизоваться по трем механизмам: в кислой и щелочной среде, а также в присутствии ферментов. В первом случае гидролиз протекает частично и избирательно. Так, например, при гидролизе ДНК происходит разрыв связей между пуриновыми основаниями и моносахаридом дезоксирибозой и образуется ДНК, в составе которой нет пуриновых оснований.
В щелочной среде нуклеиновые кислоты гидролизуются полностью, но в несколько этапов: нуклеотиды → нуклеозиды → азотистые основания, моносахарид и ортофосфорная кислота.
Ферментативный гидролиз протекает под действием специальных энзимов – нуклеаз. Происходит разрыв конкретных химических связей.
Как известно, большая часть нуклеиновых кислот в клетке связана с белком в форме нуклеопротеинов. Поступающие с пищей нуклеопротеины разрушается панкреатическими ферментами, а нуклеопротеины ткани — лизосомальными ферментами. Вначале происходит диссоциация компонентов нуклеопротеинов на белки и нуклеиновые кислоты. Этому способствует кислая среда желудка. Белки затем включаются в обмен вместе с другими белками пищи, а нуклеиновые кислоты гидролизуются нуклеазами сока железы (РНКазами и ДНКазами), с образованием смеси полинуклеотидов. Далее в процесс включаются полинуклеотидазы и фосфодиэстеразы (эндонуклеазы) кишечника Они довершают гидролиз нуклеиновых кислот до мононуклеотидов. В кишечнике, как правило, образуются 3′-фосфат нуклеотиды, а под влиянием лизосомальных полинуклеотидаз образуются биологически важные 5′- фосфат нуклеотиды. Нуклеотиды гидролизуются нуклеотидазами, с образованием нуклеозидов и Фн. Нуклеозиды, которые обычно рассматриваются как конечный продукт переваривания нуклеиновых кислот в кишечнике, всасываются.
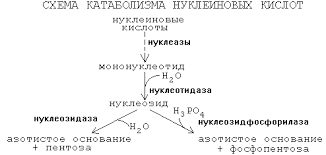
В клетках некоторых тканей, в том числе и клетках кишечника, нуклеозиды подвергаются фосфоролизу нуклеозид фосфорилазами, с образованием оснований и рибозы 1-Ф (или дезоксирибозы 1-P). Рибоза 1-Ф и рибоза 5-Ф в цитозоле находятся в равновесии и могут быть вновь использованы для синтеза нуклеотидов или вступают в неокислительную часть пенозофосфатного пути. Пуриновые и пиримидиновые основания также или распадаются далее до конечных продуктов или используются повторно для синтеза нуклеотидов. В клетке существует интенсивно обмениваемый пул рибонуклеотидов и РНК. Молекулы ДНК и пул дезоксирибонуклеотидов обменивается значительно медленнее. Тканевые пурины и пиримидины, которые не попадают в пути повторного использования, обычно распадаются и продукты их распада выделяются. Используется лишь очень небольшое количество пищевых пуринов, а основная масса поступивших с пищей пуринов распадается. Катаболизм пуринов и пиримидинов не сопровождается значительным высвобождением энергии в сравнении с обменом аминокислот, однако некоторые продукты распада выполняют определенные физиологические функции, например, конечный продукт катаболизма пуринов у человека мочевая кислота, может служить антиоксидантом, продукт катаболизма пиримидина, β– аланин используется в синтезе активных пептидов мозга и мышц. Парэнтеральное введение нуклеотидов и нуклеозидов нашло применение в исследовательской практике. Меченый 3Н –тимидин включается в синтезируемую ДНК без изменений и используется для введения метки в ДНК различных биологических объектов.
Реакции распада мочевой кислоты млекопитающих имеется фермент уратоксидаза которая превращает мочевую кислоту в более растворимый аллантоин. У человека нет такого фермента, и мочевая кислота, кото- рая образуется преимущественно в печени, выделяется почками как конечный продукт обмена пуриновых нуклеотидов. Превращение мочевой кислоты в аллантоин может протекать неферментативно. Эта реакция рассматривается как один из механизмов антиоксидантной защиты клетки у организмов, утративших способность синтезировать аскорбиновую кислоту, а мочевая кислота как важный антиоксидант, заменивший аскорбиновую кислоту. У некоторых животных аллантоин может распадаться далее до мочевины или аммиака. От нуклеотидов к основаниям. Гуаниновые нуклеотиды гидролизуются с образованием гуанозина, который подвергается фосфоролизу до гуанина и рибоза 1-Ф. Гуанин дезаминируется гуанин дезаминазой с образованием ксантина. Аденозин также можно получить по такому пути, однако внутриклеточные нуклеотидазы у человека не очень активны по отношению к AMФ. АМФ чаще дезаминируется ферментом аденилат (AMФ) дезаминазой с образованием ИМФ. Последний далее гидролизуется нуклеотидазой с образованием инозина и после фосфоролиза превращается в гипоксантин. Некоторое количество аденозина образуется из S -аденозилметионина в процессах переметилирования. Аденозин дезаминируется до инозина аденозин дезаминазой. Недостаточность аденозин дезаминазы или пуриновой нуклеозид фосфорилазы ведет к двум различным болезням иммунодефицита механизмами, которые до конца не раскрыты.
При недостаточности аденозин дезаминазы, страдают T и B-лимфоциты, а при недостаточности фосфорилазы нарушается функция T клеток, а B клетки остаются нормальными. В сентябре 1990 г была успешно применена генинженерная технология для лечения 4-летней девочки с недостаточностью аденозин дезаминазы. Катаболизм метилированных (минорных) пуринов зависит от расположения метильной группы. Если метильная группа связана с группой -NH2, она удаляется вместе с -NH2, и оставшаяся часть обменивается в дальнейшем обычным способом. Если метил связан с атомом азота гетероцикла, соединение выделяется в неизменном виде с мочой. От оснований к мочевой кислоте И адениновые и гуаниловые нуклеотиды превращаются в одно общее промежуточное соединение ксантин. Гипоксантин, возникающий из аденина, окисляется в ксантин ксантиноксидазой . Гуанин дезаминируется с образованием аммиака и ксантина. Образующийся аммиак переносится к печени в составе глутамина, где используется для синтеза мочевины. Ксантин, подобно гипоксантину, окисляется кислородом и ксантиноксидазой в мочевую кислоту с образованием перекиси водорода. У человека, мочевая кислота выделяется, а перекись водорода разрушается каталазой. Высокая активность ксантиноксидазы обнаруживается только в клетках печени и кишечника.
- Нуклеозиды и их гидролиз. Строение и гидролиз мононуклеотидов. Первичная структура нуклеиновых кислот. Фосфодиэфирная связь. ДНК и РНК: состав и гидролиз. Вторичная структура РНК и ДНК.
- Лекция № 25 нуклеиновые кислоты (45 минут)
- Лекция № 25
- Нуклеиновые кислоты (45 минут)
- 1. Компоненты нуклеиновых кислот
- а) азотистые основания
- б) углеводы
- в) фосфорная кислота
- 2. Нуклеозиды
- 3. Нуклеотиды
- а) мононуклеотиды
- б) олиго — и полинуклеотиды
- 4. Вторичная структура ДНК
- 5. Химические свойства нуклеиновых кислот.
- а) кислотный гидролиз
- б) щелочной гидролиз
- в) ферментативный гидролиз
- 1. Компоненты нуклеиновых кислот
- Азотистые основания.
- Пиримидиновые основания
- Пуриновые основания
- Углеводы.
- Фосфорная кислота.
- 2. Нуклеозиды
- Нуклеотиды
- Мононуклеотиды.
- Нуклеозидмонофосфаты.
- Нуклеозидмоноциклофосфаты.
- Нуклеозид-5′-дифосфаты (НДФ) и нуклеозид-5′-трифосфаты (НТФ).
- Олиго — и полинуклеотиды.
- Вторичная структура РНК.
- Гидролиз в кислой среде.
- Гидролиз в щелочной среде.
- Ферментативный гидролиз.
Нуклеозиды и их гидролиз. Строение и гидролиз мононуклеотидов. Первичная структура нуклеиновых кислот. Фосфодиэфирная связь. ДНК и РНК: состав и гидролиз. Вторичная структура РНК и ДНК.
N-гликозиды D-рибозы или 2-дезокси- D-рибозы и гетероциклических пуриновых и пиримидиновых оснований называются нуклеозиды. В них появляется т.н. N-гликозидная связь, возникающая на месте гликозидного гидроксила. Сахара в составе нуклеозидов входят в фуранозной форме и в виде b-аномеров.


D-рибоза 2-дезокси- D-рибоза
В зависимости от природы углеводного остатка различают рибонуклеозиды (в РНК) и дезоксирибонуклеозиды (в ДНК).
Нуклеозиды могут находиться в различных конформациях. В син— конформации гетероцикл развернут функциональными группами «внутрь» нуклеозида, в анти- конформации – «наружу». В ДНК и РНК нуклеозиды находятся в анти-конформации.
  |   |
Являясь гликозидами нуклеозиды устойчивы к гидролизу в щелочной среде, но гидролизуются в кислой среде до сахара и нуклеиновых оснований.
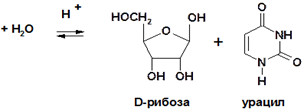
Нуклеотиды – мономеры НК. Принцип строения нуклеотидов
Нуклеотиды – это фосфорные эфиры нуклеозидов. Мономерами НК являются именно нуклеотиды. Нумерация атомов углерода в пентозе нуклеотида идет с добавлением верхнего штриха к номеру углерода, а в гетероцикле без штриха.
Фосфорная кислота образует сложноэфирные связи с первичноспиртовой группой рибозы по 5¢-углероду или со вторичноспиртовой группой при 3¢- углероде. Нуклеотиды можно рассматривать и как эфиры и как кислоты, так как они содержат остаток фосфорной кислоты с двумя кислотными центрами, При рН = 7 остаток фосфорной кислоты находится в ионизированном состоянии
Для связывания трех компонентов нуклеотидов используется сложноэфирная и N-гликозидная связи





Нуклеотиды гидролизуются как в кислой, так и в щелочной среде но с образованием разных продуктов. В щелочной среде разрушаются сложноэфирные связи, образуются нуклеозиды и анион фосфорной кислоты. В кислой среде разрушаются и сложноэфирные и гликозидные связи, образуются: фосфорная кислота, пентоза (рибоза или дезоксирибоза) и нуклеиновые основания. Поскольку нуклеотиды – мономеры НК, ДНК и РНК при гидролизе в кислой или щелочной среде образуют те же продукты, что и нуклеотиды.
В полинуклеотидных цепях нуклеиновые звенья связываются через фосфатную группу. Фосфатная группа образует две сложноэфирные связи с третьим углеродом вышестоящей рибозы (дезоксирибозы) и с пятым углеродом нижестоящей.
 Рис. Первичной структуры РНК.
Рис. Первичной структуры РНК.
Тот конец полинуклеотидной цепочки, где остается свободный первичноспиртовой гидроксил у 5¢ атом углерода пентозы, называется 5¢-конец. Там, где находится свободный гидроксил на 3¢-атоме углерода пентозы называется 3¢-конец.
Гидролиз Рнк и ДНк различен. Днк устойчивы к гидролизу в щелочной среде, а РНК гидролизируются в этих же условиях в мягких условиях. Хим. Гидролиз ДНК не применяется из-за массы побочных процессов. Более предпочтителен ферментативный катализ с участием нуклеаз , змеиного яда, в котором содержатся ферменты, расщепляющие фосфодиэфирные связи.
Под вторичной структурой понимают пространственную организацию полинуклеотидной цепи. Согласно модели Уотсона-Крика молекула ДНК состоит из двух полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали
Водородные связи образуются между аминогруппой одного основания и карбонильной группой другого -NH. O=C-, а также между амидным и иминным атомами азота -NH. N.
Первичная структура, т.е. нуклеотидная последовательность, одной цепи предопределяет первичную структуру второй цепи.
Комплементарность цепей и последовательность звеньев составляют химическую основу важнейшей функции ДНК — хранения и передачи наследственной информации.
В стабилизации молекулы ДНК наряду с водородными связями, действующими поперек спирали, большую роль играют межмолекулярные взаимодействия, направленные вдоль спирали между соседними пространственно сближенными азотистыми основаниями — стэкинг-взаимодействиями
Сильное стэкинг-взаимодействие всегда усиливает водородные связи между основаниями, способствуя уплотнению спирали. Вследствие этого молекулы воды из окружающего раствора связываются в основном с пентозофосфатным остовом ДНК, полярные группы которого находятся на поверхности спирали. При ослаблении стэкинг-взаимодействия молекулы воды, проникая внутрь спирали, конкурентно взаимодействуют с полярными группами оснований, инициируют дестабилизацию и способствуют дальнейшему распаду двойной спирали
38. Строение АТФ АДФ АМФ. Строение НАД+ и его фосфата НАДФ+.
аденозин-5′-фосфат (АМР), аденозин-5′-дифосфат (ADP)и аденозин-5′-трифосфат (ATP) Нуклеотиды, фосфорилированные в разной степени, способны к взаимопревращениям путем наращивания или отщепления фосфатных групп. Дифосфатная группа содержит одну, а трифосфатная — две ангидридные связи, называемые макроэргическими, поскольку они обладают большим запасом энергии. Необходимые для образования такой связи энергетические затраты восполняются за счет энергии, выделяемой в процессе метаболизма углеводов. При расщеплении макроэргической связи Р
О (обозначаемой волнистой линией) выделяется
32 кДж/моль. С этим связана важнейшая роль АТФ как «поставщика» энергии во всех живых клетках.
В показанных ниже взаимопревращениях АМФ, АДФ и АТФ формулы этих соединений соответствуют их неионизированному состоянию. В физиологических условиях при рН
7 фосфатные группы почти полностью ионизированы, поэтому в биохимической литературе эти и любые другие нуклеотиды записывают соответственно в виде анионов.

Нуклеозидполифосфаты в биохимических процессах. — перенос фосфатных групп. в. Все стадии гликолиза (превращения глюкозы в пируват) осуществляются только в фосфатной форме. Получение фосфатов гидроксилсодержа- щих соединений можно представить в виде общей схемы.

Так, галактоза, образующаяся при расщеплении лактозы, на начальной стадии метаболического превращения в глюкозу взаимо- действует с АТФ с образованием монофосфата.

Никотинамиднуклеотиды
Наиболее важными представителями этой группы соединений являются никотинамидадениндинуклеотид(NAD, или в русской литературе НАД) и его фосфат (NADP, или НАДФ). Эти соединения выполняют важную роль коферментов в осуществлении многихокислительно-восстановительных реакций. В соответствии с этим они могут существовать как в окисленной (НАД+, НАДФ+), так и восстановленной (НАДН, НАДФН) форме.

Структурным фрагментом НАД + и НАДФ + является никотинамидный остаток в виде пиридиниевого катиона. В составе НАДН и НАДФН этот фрагмент превращается в остаток 1,4-дигидропиридина.
В ходе биологического дегидрирования субстрат теряет два атома водорода, т. е. два протона и два электрона (2Н+, 2е) или протон и гидрид-ион (Н+ и Н — ). Кофермент НАД+ обычно рассматривается как акцептор гидрид-иона Н — (хотя окончательно не установлено, происходит ли перенос атома водорода к этому коферменту одновременно с переносом электрона или эти процессы протекают раздельно).

В результате восстановления путем присоединения гидрид-иона к НАД+ пиридиниевое кольцо переходит в 1,4-дигидропиридиновый фрагмент. Этот процесс обратим.
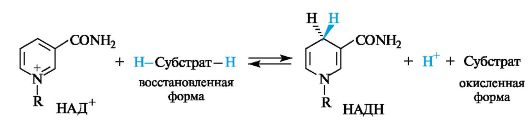
В реакции окисления ароматический пиридиниевый цикл переходит в неароматический 1,4-дигидропиридиновый цикл. В связи с потерей ароматичности возрастает энергия НАДН по сравнению с НАД + . Таким способом НАДН запасает энергию, которая затем расходуется в других биохимических процессах, требующих энергетических затрат.
Лекция № 25 нуклеиновые кислоты (45 минут)

Лекция № 25
Нуклеиновые кислоты (45 минут)
1. Компоненты нуклеиновых кислот
а) азотистые основания
б) углеводы
в) фосфорная кислота
2. Нуклеозиды
3. Нуклеотиды
а) мононуклеотиды
б) олиго — и полинуклеотиды
4. Вторичная структура ДНК
5. Химические свойства нуклеиновых кислот.
а) кислотный гидролиз
б) щелочной гидролиз
в) ферментативный гидролиз
Нуклеиновые кислоты — это водорастворимые высокомолекулярные соединения, макромолекулы которых состоят из многократно повторяющихся звеньев — нуклеотидов. Поэтому их также называют полинуклеотидами.
Впервые они были обнаружены в 1868 году швейцарским врачом Мишером в ядрах погибших лейкоцитов, что и определило их название (от лат. nucleus — ядро).
По химическому строению нуклеиновые кислоты — биологические полимеры, состоящие из остатков фосфорной кислоты, моносахарида и одного из пуриновых или пиримидиновых гетероциклических оснований. Входящие в состав нуклеиновых кислот гетероциклические соединения пуринового и пиримидинового ряда называют нуклеиновыми основаниями или просто основаниями.
В общем виде молекулу нуклеиновой кислоты можно представить следующим образом:

Фрагмент макромолекулы нуклеиновой кислоты моносахарид-основание называется нуклеозидом, а фрагмент моносахарид-основание-фосфат — нуклеотидом.
Нуклеиновые кислоты присутствуют в клетках всех живых организмов и выполняют важнейшие функции по хранению, передаче и реализации наследственной информации.
Особенность нуклеиновых кислот заключается в том, что обычно «неделимое» мономерное звено в полинуклеотидах может быть разделено на составные части: нуклеотиды и нуклеозиды, которые помимо того, что служат строительными блоками при синтезе нуклеиновых кислот, играют и вполне самостоятельную роль в процессах обмена веществ, а также участвуют в качестве коферментов в реакциях переноса остатков сахаров, аминов и других биомолекул.
1. Компоненты нуклеиновых кислот
Азотистые основания.
В состав нуклеиновых кислот входят гетероциклические основания пиримидинового ряда: урацил, тимин, цитозин и пуринового ряда: аденин и гуанин.
Для удобства используют однобуквенные обозначения (символы) гетероциклических оснований (русские или латинские):
Пиримидиновые основания

Пуриновые основания

Нумерация атомов производится в указанном порядке. Отметим также, что в физиологических условиях нуклеиновые основания существуют только в лактамной и аминной формах. Во всех формах гетероциклы сохраняют ароматичность и имеют плоское строение, благодаря чему отличаются высокой термодинамической стабильностью.
Углеводы.
Углеводным компонентом нуклеиновых кислот являются пентозы: D-рибоза или 2-дезокси-D-рибоза. Обе пентозы всегда находятся в фуранозной форме и имеют β-конфигурацию аномерного атома С-1′ (в формулах нуклеотидов и нуклеозидов атомы углерода моносахаридов нумеруются цифрой со штрихом):

Фосфорная кислота.
Остатки фосфорной кислоты (H3PO4) входят в состав нуклеотидов и этерифицируют D-рибозу и 2-дезокси-D-рибозу в положениях 3′ и 5′. В этой связи нуклеотиды и полинуклеотиды можно рассматривать, с одной стороны, как сложные эфиры нуклеозидов (фосфаты), а с другой — как кислоты.
Остатки моносахаридов и фосфорной кислоты выполняют структурную функцию и являются носителями гетероциклических оснований.
2. Нуклеозиды
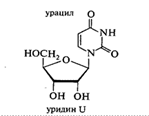
Нуклеозиды состоят из гетероциклических оснований, связанных с моносахаридом N-гликозидной связью. В образовании этой связи участвуют аномерный атом углерода моносахарида и атомы азота N-1 (у пиримидинового основания) и N-9 (у пуринового основания):

В зависимости от углеводного остатка нуклеозиды делятся на рибонуклеозиды и дезоксирибонуклеозиды. Номенклатурные названия нуклеозидов строятся аналогично названиям гликозидов, например, β-аденинрибофуранозид и т. д. Однако чаще используются названия с суффиксами — идин у пиримидиновых и — озин у пуриновых нуклеозидов и приставкой дезокси — у дезоксирибонуклеозидов:
цитозин + рибоза = цитидин;
цитозин + дезоксирибоза = дезоксицитидин;
аденин + рибоза = аденозин;
аденин + дезоксирибоза = дезоксиаденозин.
Исключением является название тимидин, используемое для дезокситимидина, входящего в состав ДНК.
Кроме того, нуклеозиды обозначают символами, производимыми от входящих в них оснований. В случае дезоксирибонуклеозидов перед символом ставят строчную букву д (лат. d). Символами удобно пользоваться при обозначении первичной структуры ДНК или РНК. Если речь идет только о ДНК, букву d обычно пропускают.








Нуклеозиды значительно лучше растворимы в воде, чем исходные азотистые основания. Подобно всем гликозидам, нуклеозиды устойчивы к действию щелочей, но при нагревании легко подвергаются кислотному гидролизу с разрывом гликозидной связи и образованием основания и пентозы:

Пиримидиновые нуклеозиды значительно более устойчивы к гидролизу, чем пуриновые. В условиях in vivo гидролиз обоих типов нуклеозидов осуществляется при помощи специфических ферментов, называемых нуклеозидазами.
Нуклеотиды
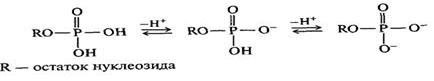
Нуклеотиды — это производные нуклеозидов, которые образуются в результате этерификации пентозного фрагмента фосфорной кислотой. В зависимости от строения пентозы различают рибонуклеотиды (мономерные звенья РНК) и дезоксирибинуклеотиды (мономерные звенья ДНК).
В 2-дезоксирибозе имеются только два положения, по которым может образовываться сложноэфирная связь — а именно 3′- и 5′-положения. В случае рибонуклеотидов фосфатная группа может находиться в положениях 2′, 3′ и 5′. В биологических системах встречаются нуклеотиды всех типов, однако наиболее распространены нуклеозид-5′-фосфаты. За счет фосфатного остатка нуклеотиды проявляют свойства кислот и в физиологических условиях (pH ≈ 7) находятся в практически полностью ионизированном состоянии.
Мононуклеотиды.
Мононуклеотиды — это повторяющиеся мономерные единицы олигонуклеотидов и полинуклеотидов.
Нуклеозидмонофосфаты.
Нуклеозидмонофосфаты содержат только один остаток фосфорной кислоты. Например:

Для мононуклеотидов также используют два вида названий (табл. 3.), отражающих их принадлежность как к сложным эфирам, так и к кислотам. В первом случае название включает наименование нуклеозида с указанием положения в нем фосфатного остатка: аденозин-5′-фосфат*, уридин-5′-фосфат и т. д. Широко используются и тривиальные названия, имеющие окончание «-овая кислота», например, 5′-Уридиловая кислота.
Для удобства введены сокращения: трехбуквенные — АМФ (лат. AMP), УМФ (лат. UMP) и т. д. и однобуквенные — pA, pU и т. д. В биохимической литературе встречаются как русские, так и латинские обозначения, однако более широко используются латинские. Сокращения AMP, GMF и т. д. всегда относятся к 5′-нуклеотидам. У других, например, 3′-производных в сокращенном названии указывается положение фосфатной группы: 3′-AMP, 3′-GMP и т. д.
* вместо «монофосфат» часто пишут просто «фосфат».
В однобуквенных кодах 5′-фосфаты записываются с добавлением латинской буквы «p» перед символом нуклеозида (pA), 3′-фосфаты — после символа нуклеозида (Ap).
Важнейшие нуклеотиды, входящие в состав нуклеиновых кислот.
Входящие в состав РНК
Входящие в состав ДНК
Сложноэфирные связи с фосфатными остатками в мононуклеотидах относительно устойчивы к кислотному гидролизу. Однако фермент 5′-нуклеотидаза способен отщеплять фосфатную группу в положении 5′, не затрагивая при этом N-гликозидную связь.
Нуклеозидмоноциклофосфаты.
Существуют нуклеотиды, в которых фосфорная кислота одновременно этерифицирует две гидроксильные группы пентозного остатка с образованием устойчивого шестичленного цикла. В этой связи в их названиях используют приставку цикло-, например, циклоаденозин-монофосфат (cAMP) или циклогуанозинмонофосфат (cGMP). Эти два нуклеозидциклофосфата присутствуют практически во всех клетках живых организмов:

Биологическая роль циклических нуклеотидов была открыта сравнительно недавно. Так, аденозин-3′, 5′-циклофосфат выполняет роль внутриклеточного «посредника» в процессах, индуцируемых гормоном адреналином.
Нуклеозид-5′-дифосфаты (НДФ) и нуклеозид-5′-трифосфаты (НТФ).
Во всех тканях организма в свободном состоянии содержатся не только моно-, но и ди-, а также трифосфаты нуклеозидов. Наиболее известными являются аденозин-5′-дифосфат (АДФ) аденозин-5′-трифосфат (АТФ).
Нуклеотиды, фосфорилированные в разной степени, способны к взаимопревращениям путем наращивания или отщепления фосфатных групп. Дифосфатная группа содержит одну, а трифосфатная — две ангидридные связи, называемые макроэргическими, поскольку они обладают большим запасом энергии. При расщеплении макроэргической связи Р — О выделяется энергия, приблизительно равная 32 кДж/моль. Этим объясняется важнейшая роль АТФ как «поставщика» энергии во всех живых клетках.
Ангидридные связи могут гидролизоваться либо специфичными ферментами, либо при кипячении в присутствии HCl. Ни сложноэфирная связь, ни N-гликозидная при этом не расщепляются.
Взаимные превращения АМФ, АДФ и АТФ.
Олиго — и полинуклеотиды.
Олигонуклеотидами называют соединения, в которых несколько нуклеозидов (до 20) соединены между собой фосфодиэфирными связями. Нуклеозид со свободной 5′-OH-групой называется
5′-концевым, а со свободной 3′-OH-групой — 3′-концевым.
Полинуклеотиды представляют собой линейные гетерополимеры, также состоящие из последовательности мононуклеозидных единиц, соединенных фосфатным мостиком. Полинуклеотиды, состоящие из ковалентно связанных между собой остатков дезоксирибонуклеозидов, называются дезоксирибонуклеиновыми кислотами (ДНК), а полинуклеотиды, состоящие из остатков рибонуклеозидов — рибонуклеиновыми кислотами (РНК). В обоих случаях мононуклеозиды связаны между собой при помощи фосфодиэфирных мостиков, соединяющих 3′-положение одного мононуклеотида с 5′-положением его соседа.
Нуклеиновые кислоты также различаются и по составу входящих в них гетероциклических оснований: урацил входит только в состав РНК, а тимин — только в состав ДНК. Аденин, цитозин и гуанин являются постоянными компонентами полинуклеотидов.
Молекулярная масса ДНК достигает десятков миллионов. Это самые длинные из известных макромолекул. Значительно меньше молекулярная масса РНК (от нескольких сотен до десятков тысяч). ДНК содержатся в основном в ядрах клеток, РНК — в рибосомах и протоплазме клеток.
Первичная структура нуклеиновых кислот.
Первичная структура нуклеиновых кислот — это нуклеотидный состав и определенная последовательность нуклеотидных звеньев в полимерной цепи..
Для записи первичной структуры ДНК и РНК используют однобуквенные обозначения нуклеозидов. Приведенные фрагменты ДНК и РНК записывают соответственно: d(A-C-G-T) и (A-C-G-U). Часто букву d опускают, если речь идет о ДНК.


Первичная структура цепи ДНК и РНК.
Нуклеотидный состав ДНК различного происхождения находится в соответствии с правилами Э. Чаргаффа:
1) Все ДНК независимо от их происхождения содержат одинаковое число пуриновых и пиримидиновых оснований. Следовательно, в любой ДНК на каждый пуриновый нуклеотид приходится один пиримидиновый.
2) Любая ДНК всегда содержит в равных количествах попарно аденин и тимин, гуанин и цитозин, что обычно обозначают как А = Т и G = C.
3) Количество оснований, содержащих аминогруппы в положении 4 пиримидинового ядра и 6 пуринового (цитозин и аденин), равно количеству оснований, содержащих оксо-группу в тех же положениях (гуанин и тимин), т. е. A + C=G + T.
Наряду с этим было установлено, что для каждого типа ДНК суммарное содержание гуанина и цитозина не равно суммарному содержанию аденина и тимина, т. е. что (G + C)/(A + T), как правило, отличается от единицы (может быть как больше, так и меньше ее). По этому признаку различают два основных типа ДНК: А-Т-тип с преимущественным содержанием аденина и тимина и G-C-тип с преимущественным содержанием гуанина и цитозина.
Для РНК правила Чаргаффа либо не выполняются, либо выполняются с большим приближением.
Вторичная структура нуклеиновых кислот.
Под вторичной структурой нуклеиновых кислот понимают пространственно упорядоченные формы полинуклеотидных цепей.
Вторичная структура ДНК представляет собой две параллельные неразветвленные полинуклеотидные цепи, закрученные вокруг общей оси в двойную спираль.

Вторичная структура ДНК.
Такая пространственная структура удерживается множеством водородных связей, образуемых азотистыми основаниями, направленными внутрь спирали. Эти основания составляют комплементарные пары (от лат. complementum — дополнение). Образование водородных связей между комплементарными парами оснований обусловлено их пространственным соответствием. Так, пиримидиновое основание комплементарно пуриновому:

Водородные связи между другими парами оснований не возникают по пространственным причинам. Таким образом, в ДНК:
ТИМИН (Т) комплементарен АДЕНИНУ (А)
ЦИТОЗИН (C) комплементарен ГУАНИНУ (G)
Комплементарность оснований определяет комплементарность цепей в молекулах ДНК. Водородные связи возникают между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи. Иными словами, пуриновым основаниям аденину и гуанину в одной цепи будут соответствовать пиримидиновые основания тимин и цитозин в другой цепи.
Таким образом, полинуклеотидные цепи, образующие двойную спираль, не идентичны, но комплементарны между собой:
В сдвоенной молекуле ДНК суммарно число групп А равно числу групп Т, а число групп G — числу C.
Комплементарность полинуклеотидных цепей служит химической основой главной функции ДНК — хранения и передачи наследственных признаков. Способность ДНК не только хранить, но и использовать генетическую информацию определяется следующими ее свойствами:
1) Молекулы ДНК способны к репликации (удвоению), т. е. могут обеспечить возможность синтеза других молекул ДНК, идентичных исходным, поскольку последовательность оснований в одной из цепей двойной спирали контролирует их расположение в другой цепи;
2) Молекулы ДНК могут совершенно точным и определенным образом направлять синтез белков, специфичных для организмов данного вида.
Вторичная структура РНК.
В отличие от ДНК, молекулы РНК состоят из одной полинуклеотидной цепи и не имеют строго определенной пространственной формы (рис. 3.), т. к. вторичная структура РНК зависит от их биологических функций. Полимерная цепь РНК приблизительно в десять раз короче, чем у ДНК.
Комплементарными парами в РНК являются цитозин-гуанин и урацил-аденин.

Вторичная структура РНК.
Основная роль РНК — непосредственное участие в биосинтезе белка. Известны три вида клеточных РНК, которые отличаются по местоположению в клетке, составу, размерам и свойствам, определяющим их специфическую роль в образовании белковых макромолекул:
1) Информационные (матричные) РНК передают закодированную в ДНК информацию о структуре белка от ядра клетки к рибосомам, где и осуществляется синтез белка;
2) Транспортные РНК собирают аминокислоты в цитоплазме клетки и переносят их в рибосому; молекулы РНК этого типа «узнают» по соответствующим участкам цепи информационной РНК, какие аминокислоты должны участвовать в синтезе белка;
3) Рибосомные РНК обеспечивают синтез белка определенного строения, считывая информацию с информационной (матричной) РНК.
Химические свойства нуклеиновых кислот.
ДНК и РНК имеют много общих химических и физических свойств. Так, нуклеиновые кислоты хорошо растворимы в воде и плохо растворяются в водных растворах кислот. Существенные различия ДНК и РНК связаны, в основном, с их отношением к гидролизу.
Гидролиз в кислой среде.
Мягкий кислотный гидролиз ДНК оказывает весьма избирательное действие: он приводит к расщеплению N-гликозидных связей между пуриновыми основаниями и дезоксирибозой, связи пиримидин-дезоксирибоза при этом не затрагиваются. В результате образуется ДНК, лишенная пуриновых оснований. Гидролиз РНК, проводимый в аналогичных условиях, приводит к образованию пуриновых оснований и пиримидиновых нуклеозид-2′(3′)-фосфатов.
Кислотный гидролиз в жестких условиях, приводит к разрыву всех N-гликозидных связей как ДНК, так и РНК и образованию смеси пуриновых и пиримидиновых оснований.
Гидролиз в щелочной среде.
В щелочной РНК легко гидролизуются до нуклеотидов, которые в свою очередь, расщепляются с образованием нуклеозидов и остатков фосфорной кислоты. ДНК, в отличие от РНК, устойчивы к щелочному гидролизу.
Ферментативный гидролиз.
Гидролиз ДНК и РНК также протекает и при участии специфических ферментов — нуклеаз. Эти ферменты избирательно действуют на 3′- и 5′-сложноэфирные связи. Так, фосфодиэстераза, выделенная из яда змей, расщепляет все 3′-связи как в ДНК, так и в РНК с образованием нуклеозид-5′-фосфатов. Фосфодиэстераза, выделенная из селезенки быка, наоборот, гидролизует только 5′-связи и высвобождает только нуклеозид-3′-фосфаты. Известны дезокси-рибонуклеазы, расщепляющие связи только между определенными парами мононуклеотидов — их используют для направленного гидролиза РНК.
Гидролиз ДНК И РНК проводят, в основном, для установления их нуклеотидного состава. Далее выделенные смеси анализируют с использованием хроматографических и спектральных методов анализа, а также электрофореза.