Принцип метода.Фосфатидилхолины содержат в своем составе аминоспирт холин, который является сильным основанием, полностью диссоциирует в водной среде. Фосфатидилхолин имеет следующее строение:

Реактивы:лецитин;едкий натрий, 10% раствор.
Ход работы:В фарфоровую чашку наливают примерно 1 мл спиртового раствора лецитина и добавляют равное количество 10% раствора едкого натра. Кипятят в течение 2-3 мин, помешивая стеклянной палочкой. Образования триметиламина обнаруживается по запаху селедочного рассола. Результат опыта и уравнение реакции записывают.
Качественная реакция на холестерин (реакция Шиффа).
Реактивы:холестерин, хлороформный раствор; концентрированная серная кислота.

Ход работы:В пробирку наливают 1 мл хлороформного раствора холестерина и добавляют по стенке 1 мл конц. серной кислоты. На границе двух жидкостей появляется кольцо красного цвета (необходимо соблюдать осторожность при приливании серной кислоты, иначе, жидкость может выплеснуться из пробирки!). Результат опыта и уравнение реакции записывают.
Вопросы для самоконтроля:
1. Напишите строение смешанного глицерида, образованного глицерином, а также стеариновой, олеиновой и линоленовой кислотами.
2. С помощью каких реакций можно определить составные части лецитина?
3. Какова химическая природа холестерина и холестеридов?
4. Что такое омыление жиров?
5. Как можно определить составные части жира?
ЛАБОРАТОРНАЯ РАБОТА № 9
УГЛЕВОДЫ.
Теоретическая часть.
Углеводами называются альдегиды или кетоны многоатомных спиртов и полимеры этих соединений. Общая формула углеводов Cn (H2O) n. Обширная группа углеводов может быть подразделена на три класса: моносахариды, олигосахариды и полисахариды.
Моносахариды – простейшие углеводы – представляют собой соединения, характеризующиеся смешанными функциями и содержанием одновременно гидроксильных и карбонильных групп. Образуют таутомерные формы циклических полуацеталей. В зависимости от характера группы различают альдозы (полигидроксиальдегиды) и кетозы (полигидроксикетоны). Наиболее распространенными из моносахаридов в физиологическом отношении являются гексозы. Среди них глюкоза (виноградный сахар, декстроза) и фруктоза (плодовый сахар, левулоза).
 |
Свойства моносахаридов связаны с их структурным строением, т.е. наличием в их молекуле спиртовых и альдегидных групп. Как спирты они образуют алкоголяты. Как альдегиды они способны к реакции полимеризации и реакциям восстановления металлов и окисления углевода.
Олигосахариды (от греч. «олигос» — немногий) – полные ацетали, образуются с выделением воды из двух или нескольких (до десяти) молекул моносахаридов. По своему строению олигосахариды являются гликозидами. Олигосахариды сравнительно легко гидролизуются с образованием соответствующих моносахаридов. Они дают истинные растворы, обладают, подобно маннозам, сладким вкусом и обычно хорошо кристаллизуются. В зависимости от числа остатков моносахаридов, входящих в молекулы олигосахаридов, последние делят на дисахариды, трисахариды и т.д. Широко распространенной группой являются дисахариды.
Дисахариды, построенные из молекул гексоз, встречаются в организмах растений (например, сахароза, мальтоза) и животных (лактоза, мальтоза). Дисахариды построены по типу гликозидов. Это означает, что при образовании дисахарида одна молекула моносахарида образует связь с другой молекулой за счет своего гликозидного гидроксила. Вторая молекула моносахарида участвует в образовании связи двумя путями: 1) спиртовым гидроксилом; 2) гликозидным гидроксилом. В первом случае в молекуле дисахарида остается свободным гликозидный гидроксил, благодаря чему эти дисахариды обладают восстанавливающими свойствами (мальтоза, лактоза, целлобиоза). Во втором случае в молекуле дисахарида нет свободного гликозидного гидроксила, вследствие чего он лишен восстанавливающих свойств (сахароза, трегалоза).
Полисахариды являются полимерными соединениями, построенными по тому же принципу, что и олигосахариды. Они имеют высокий молекулярный вес. Нерастворимы в воде или дают коллоидные растворы, аморфны. Полисахариды не обладают сладким вкусом. В результате гидролиза в качестве промежуточных продуктов образуют олигосахариды, а в качестве конечных продуктов моносахариды. В организмах растений очень распространены крахмал и клетчатка, состоящие из молекул гексоз, реже встречается инулин, построенный из молекул фруктозы в организмах животных представлен полисахарид гликоген, близкий по своему строению к крахмалу. В полисахаридах гликозидный гидроксил одной молекулы моносахарида соединен со спиртовым гидроксилом второй молекулы через гликозидную связь. Полисахариды имеют свободный тликозидный гидроксил на большое число молекул моносахарида, поэтому они практически не проявляют восстанавливающих свойств.
Видео:ГИДРОЛИЗ СОЛЕЙ | 9 класс | Кратко и понятноСкачать

Фосфолипиды: фосфатидилэтаноламины, фосфатидилсерины, фосфатидилхолины, фосфатидилинозитолы: состав, биологическая роль. Реакции гидролиза и образования
Фосфоглицериды – это количественно преобладающий класс мембранных липидов. Состоят из четырех компонентов, соединенных сложноэфирными связями:
- Глицерин
- 2 остатка ВЖК
- Фосфорная кислота
- Аминоспирт (азотистое основание):
- Серин: ОН-СН2-СН-СОО —
 │
│
 NH3 +
NH3 +
·  Этаноламин: ОН-СН2-СН2— NH3 +
Этаноламин: ОН-СН2-СН2— NH3 +
 СН3 (метилирование)
СН3 (метилирование)
· 

 Инозит: ОН ОН ОН
Инозит: ОН ОН ОН
 | |
Представим структуру фосфоглицерида в общем виде:
 СН2 – О – СО – R
СН2 – О – СО – R
 │ гидрофобная часть
│ гидрофобная часть

 СН – О – СО – R
СН – О – СО – R
 ║ полярная головка
║ полярная головка
СН2 – О – Р – О – аминоспирт
Рис. 8. Структура фосфоглицерида в общем виде
Структурная особенность: фосфолипиды – амфипатические соединения, имеют «полярную головку» (фосфорную кислоту и аминоспирт) и гидрофобную часть (радикалы ВЖК).
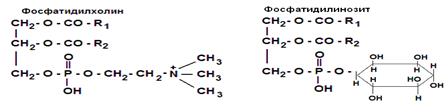
Часть структуры фосфоглицерида без 4-го компонента называется фосфатидной кислотой, её остаток – фосфатидил. Фосфоглицериды являются производными фосфатидной кислоты, этерифицированной каким-либо из аминоспиртов, и имеют соответствующее название: фосфатидилхолином фосфатидилэтаноламином, фосфатидилсерином, фосфатидилинозитолом (рис.9).
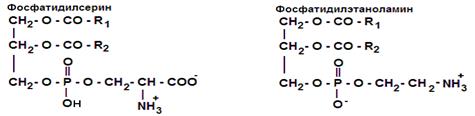
Рис. 9. Строение фосфоглицеридов
В первом положении, как правило, находятся насыщенные жирные кислоты, а во втором — ненасыщенные — (олеиновая кислота) или полиненасыщенные жирные кислоты (ПЖК).
Функции фосфолипидов:
1. ФЛ — амфифильные соединения, являются основными структурными компонентами липидного бислоя биологических мембран.
2. ФЛ, этерифицированные по второй позиции полиненасыщенными ЖК, окружают мембранные интегральные белки, создавая специфические микроусловия («гидрофобные воротнички»). Такие ФЛ получили названия «аннулярных» (окружающих).
3. Монослой липопротеинов содержит преимущественно фосфатидилхолины, которые обеспечивают некоторую гидрофильность липопротеинов.
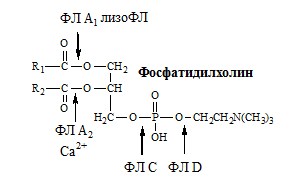
Гидролиз фосфолипидовосуществляют фосфолипазы А1, А2, С, Д, выделяемые с панкреатическим соком, которые расщепляют сложно-эфирные связи фосфолипидов в строго определенных положениях:
Фосфолипаза А1 гидролизует эфирную связь в положении 1.
Фосфолипаза А2 гидролизует эфирную связь в положении 2.
Фосфолипаза С вызывает гидролиз связи между фосфорной кислотой и глицерином.
Фосфолипаза Д расщепляет эфирную связь между аминоспиртом и фосфорной кислотой.
 |
Рис. 10. Места ферментативной атаки различными фосфолипазами (в качестве примера изображена структура фосфатидилхолина – лецитина)
При полном гидролизе фосфатидилхолина образуются глицерин, 2 ВЖК, Н3РО4, аминоспирт.
Видео:Гидролиз солей. 1 часть. 11 класс.Скачать

Липиды. Жирные кислоты. Холестерин. Метаболизм, биохимия.
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
Липиды. Переваривание и всасывание. Транспорт липидов в организме.
Обмен липопротеидов. Дислипопротеидемии.
Липиды – это разнообразная по строению группа органических веществ, которые объединены общим свойством – растворимостью в неполярных растворителях.
ЛИПИДЫ. КЛАССИФИКАЦИЯ
Липиды по способности к гидролизу делят на омыляемые (двух и более компонентные) и неомыляемые (однокомпонентные).
Омыляемые липиды в щелочной среде гидролизуются с образованием мыл, они содержат в своем в составе жирные кислоты и спирты глицерин (глицеролипиды) или сфингозин (сфинголипиды). По количеству компонентов омыляемые липиды делятся на простые (состоят из 2 классов соединений) и сложные (состоят из 3 и более классов).
К простым липидам относятся:
- 1) воска (сложный эфир высшего одноатомного спирта и жирной кислоты);
- 2) триацилглицериды, диацилглицериды, моноацилглицериды (сложный эфир глицерина и жирных кислот). У человека весом в 70 кг ТГ около 10 кг.
- 3) церамиды (сложный эфир сфингозина и жирной кислоты С18-26) – лежат в основе сфинголипидов;
К сложным липидам относятся:
1) фосфолипиды (содержат фосфорную кислоту):
- а) фосфоглицеролипиды (сложный эфир глицерина и 2 жирных кислот, содержит фосфорную кислоту и аминоспирт) – фосфатидилсерин, фосфатидилэтаноламин, фосфатидилхолин, фосфатидилинозитол, фосфатидилглицерол;
- б) кардиолипины (2 фосфатидные кислоты, соединенные через глицерин);
в) плазмалогены (сложный эфир глицерина и жирной кислоты, содержит ненасыщенный одноатомный высший спирт, фосфорную кислоту и аминоспирт) – фосфатидальэтаноламины, фосфатидальсерины, фосфатидальхолины; - г) сфингомиелины (сложный эфир сфингозина и жирной кислоты С18-26, содержит фосфорную кислоту и аминоспирт – холин);
2) гликолипиды (производные сфингозина, содержащие углеводы):
- а) цереброзиды (сложный эфир сфингозина и жирной кислоты С18-26, содержит гексозу: глюкозу или галактозу);
- б) сульфатиды (сложный эфир сфингозина и жирной кислоты С18-26, содержит гексозу (глюкозу или галактозу) к которой присоединена в 3 положение серная кислота). Много в белом веществе;
- в) ганглиозиды (сложный эфир сфингозина и жирной кислоты С18-26, содержит олигосахарид из гексоз и сиаловых кислот). Находятся в ганглиозных клетках;
К неомыляемым липидам относят:
- 1. стероиды;
- 2. жирные кислоты (структурный компонент омыляемых липидов),
- 3. витамины А, Д, Е, К;
- 4. терпены (углеводороды, спирты, альдегиды и кетоны с несколькими звеньями изопрена).
ЛИПИДЫ. БИОЛОГИЧЕСКИЕ ФУНКЦИИ
В организме липиды выполняют разнообразные функции:
- 1) Структурная. Сложные липиды и холестерин амфифильны, они образуют все клеточные мембраны; фосфолипиды выстилают поверхность альвеол, образуют оболочку липопротеинов. Сфингомиелины, плазмалогены, гликолипиды образуют миелиновые оболочки и другие мембраны нервных тканей.
- 2) Энергетическая. В организме до 33% всей энергии АТФ образуется за счет окисления липидов;
- 3) Антиоксидантная. Витамины А, Д, Е, К препятсвуют СРО;
- 4) Запасающая. ТГ являются формой хранения жирных кислот;
- 5) Защитная. ТГ, в составе жировой ткани, обеспечивают теплоизоляционную и механическую защиту тканей. Воска образуют защитную смазку на коже человека;
- 6) Регуляторная. Фосфотидилинозитолы являются внутриклеточными посредниками в действии гормонов (инозитолтрифосфатная система). Из полиненасыщенных жирных кислот образуются эйкозаноиды (лейкотриены, тромбоксаны, простагландины, простациклины), вещества, регулирующие иммуногенез, гемостаз, неспецифическую резистентность организма, воспалительные, аллергические, пролиферативные реакции. Из холестерина образуются стероидные гормоны: половые, кортикоиды, кальцитриол;
- 7) Пищеварительная. Из холестерина синтезируются желчные кислоты. Желчные кислоты, фосфолипиды, холестерин обеспечивают эмульгирование и всасывание липидов;
- 8) Информационная. Ганглиозиды обеспечивают межклеточные контакты.
Источником липидов в организме являются синтетические процессы и пища. Некоторые липиды в организме не синтезируются (полиненасыщенные жирные кислоты – витамин F, витамины А, Д, Е, К), они являются незаменимыми и поступают в организм только с пищей.
ПРИНЦИПЫ НОРМИРОВАНИЯ ЛИПИДОВ В ПИТАНИИ
В сутки человеку требуется потреблять 80-100г липидов, из них 25-30г растительного масла, 30-50г сливочного масла и 20-30г жира животного происхождения.
Потребность в пищевых липидах зависит от возраста. Новорожденным до 3 месяцев требуется 6,5 г/кг липидов, детям до 6 месяцев – 6 г/кг, детям после 6 месяцев – 5,5 г/кг, взрослым – 1,4 г/кг, пожилым – 0,5 г/кг. Причины: 1). основным источником энергии для детей грудного возраста являются липиды, а для взрослых людей – глюкоза. 2). Энергозатраты с возрастом снижаются.
Потребность в липидах увеличивается на холоде, при физических нагрузках, в период выздоровления и при беременности.
С пищей в норме поступает около 85-90г ТГ, 1г ФЛ, 0,3—0,5 г ХС (в основном в виде эфиров). Растительные масла содержат много полиеновых незаменимых (линолевая до 60%, линоленовая) жирных кислот, фосфолипидов (удаляются при рафинировании). Сливочное масло содержит много витаминов А, Д, Е.
Все природные липиды хорошо перевариваются, масла усваиваются лучше жиров. При смешанном питании сливочное масло усваивается на 93-98%, свиной жир – на 96-98%, говяжий жир – на 80-94%, подсолнечное масло – на 86-90%. Длительная тепловая обработка (> 30 мин) разрушает полезные липиды, при этом образуются токсические продукты окисления жирных кислот и канцерогенные вещества.
При недостаточном поступлении липидов с пищей снижается иммунитет, снижается продукция стероидных гормонов, нарушается половая функция. При дефиците линолевой кислоты развивается тромбоз сосудов и увеличивается риск раковых заболеваний. При избытке липидов в пище развивается атеросклероз и увеличивается риск рака молочной железы и толстой кишки.
ПЕРЕВАРИВАНИЕ ЛИПИДОВ
Переваривание – это гидролиз пищевых веществ до их ассимилируемых форм.
Лишь 40-50% пищевых липидов расщепляется полностью, от 3% до 10% пищевых липидов всасываются в неизмененном виде.
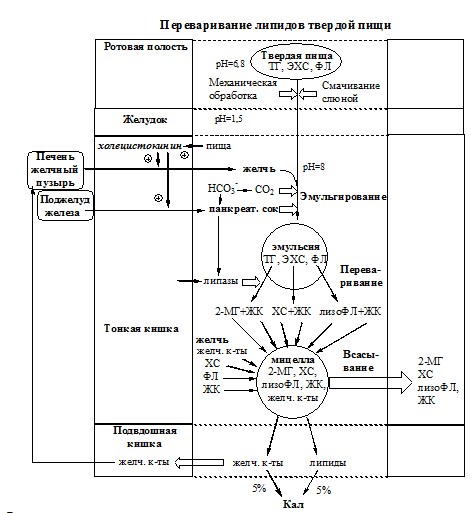
Так как липиды не растворимы в воде, их переваривание и всасывание имеет свои особенности и протекает в несколько стадий:
- 1) Липиды твердой пищи при механическом воздействии и под влиянием ПАВ желчи смешиваются с пищеварительными соками с образованием эмульсии (масло в воде). Образование эмульсии необходимо для увеличения площади действия ферментов, т.к. они работают только в водной фазе. Липиды жидкой пищи (молоко, бульон и т.д.) поступают в организм сразу в виде эмульсии;
- 2) Под действием липаз пищеварительных соков происходит гидролиз липидов эмульсии с образованием водорастворимых веществ и более простых липидов;
- 3) Выделенные из эмульсии водорастворимые вещества всасываются и поступают в кровь. Выделенные из эмульсии более простые липиды, соединяясь с компонентами желчи, образуют мицеллы;
- 4) Мицеллы обеспечивают всасывание липидов в клетки эндотелия кишечника.
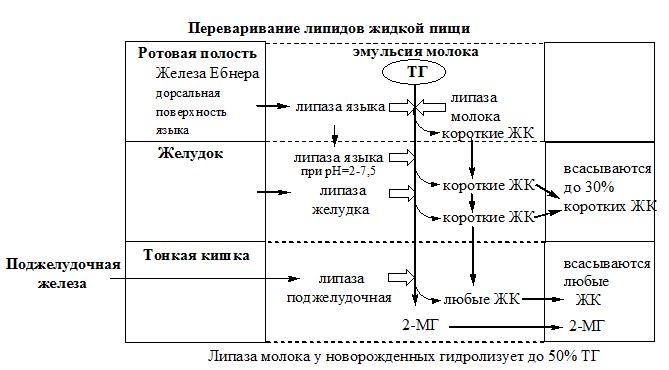
Ротовая полость
В ротовой полости происходит механическое измельчение твердой пищи и смачивание ее слюной (рН=6,8).
У грудных детей здесь начинается гидролиз ТГ с короткими и средними жирными кислотами, которые поступают с жидкой пищей в виде эмульсии. Гидролиз осуществляет лингвальная триглицеридлипаза («липаза языка», ТГЛ), которую секретируют железы Эбнера, находящиеся на дорсальной поверхности языка.
Желудок
Так как «липаза языка» действует в диапазоне 2-7,5 рН, она может функционировать в желудке в течение 1-2 часов, расщепляя до 30% триглицеридов с короткими жирными кислотами. У грудных детей и детей младшего возраста она активно гидролизует ТГ молока, которые содержат в основном жирные кислоты с короткой и средней длиной цепей (4—12 С). У взрослых людей вклад «липазы языка» в переваривание ТГ незначителен.
В главных клетках желудка вырабатывается желудочная липаза, которая активна при нейтральном значении рН, характерном для желудочного сока детей грудного и младшего возраста, и не активна у взрослых (рН желудочного сока
1,5). Эта липаза гидролизует ТГ, отщепляя, в основном, жирные кислоты у третьего атома углерода глицерола. Образующиеся в желудке ЖК и МГ далее участвуют в эмульгировании липидов в двенадцатиперстной кишке.
Тонкая кишка
Основной процесс переваривания липидов происходит в тонкой кишке.
1. Эмульгирование липидов (смешивание липидов с водой) происходит в тонкой кишке под действием желчи. Желчь синтезируется в печени, концентрируется в желчном пузыре и после приёма жирной пищи выделяется в просвет двенадцатиперстной кишки (500-1500 мл/сут).
Жёлчь это вязкая жёлто-зелёная жидкость, имеет рН=7,3-8.0, содержит Н2О – 87-97%, органические вещества (желчные кислоты – 310 ммоль/л (10,3-91,4 г/л), жирные кислоты – 1,4-3,2 г/л, пигменты желчные – 3,2 ммоль/л (5,3-9,8 г/л), холестерин – 25 ммоль/л (0,6-2,6) г/л, фосфолипиды – 8 ммоль/л) и минеральные компоненты (натрий 130-145 ммоль/л, хлор 75-100 ммоль/л, НСО3- 10-28 ммоль/л, калий 5-9 ммоль/л). Нарушение соотношение компонентов желчи приводит к образованию камней.
Жёлчные кислоты (производные холановой кислоты) синтезируются в печени из холестерина (холиевая, и хенодезоксихолиевая кислоты) и образуются в кишечнике (дезоксихолиевая, литохолиевая, и д.р. около 20) из холиевой и хенодезоксихолиевой кислот под действием микроорганизмов. В желчи желчные кислоты присутствуют в основном в виде конъюгатов с глицином (66-80%) и таурином (20-34%), образуя парные желчные кислоты: таурохолевую, гликохолевую и д.р.
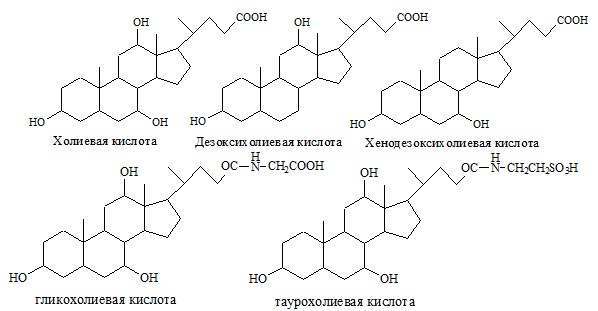
Соли жёлчных кислот, мыла, фосфолипиды, белки и щелочная среда желчи действуют как детергенты (ПАВ), они снижают поверхностное натяжение липидных капель, в результате крупные капли распадаются на множество мелких, т.е. происходит эмульгирование. Эмульгированию также способствует перистальтика кишечника и выделяющийся, при взаимодействии химуса и бикарбонатов СО2: Н+ + НСО3- → Н2СО3 → Н2О + ↑СО2.
2. Гидролиз триглицеридов осуществляет панкреатическая липаза. Ее оптимум рН=8, она гидролизует ТГ преимущественно в положениях 1 и 3, с образованием 2 свободных жирных кислот и 2-моноацилглицерола (2-МГ). 2-МГ является хорошим эмульгатором.
28% 2-МГ под действием изомеразы превращается в 1-МГ. Большая часть 1-МГ гидролизуется панкреатической липазой до глицерина и жирной кислоты.
В поджелудочной железе панкреатическая липаза синтезируется вместе с белком колипазой. Колипаза образуется в неактивном виде и в кишечнике активируется трипсином путем частичного протеолиза. Колипаза своим гидрофобным доменом связывается с поверхностью липидной капли, а гидрофильным способствует максимальному приближению активного центра панкреатической липазы к ТГ, что ускоряет их гидролиз.
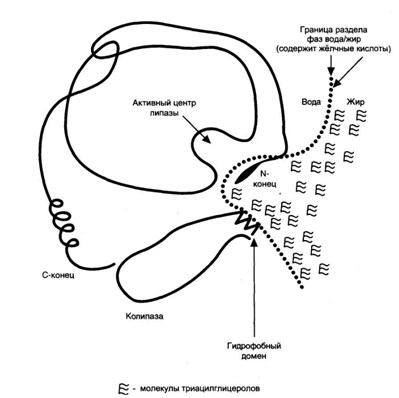
3. Гидролиз лецитина происходит с участием фосфолипаз (ФЛ): А1, А2, С, D и лизофосфолипазы (лизоФЛ).
В результате действия этих четырех ферментов фосфолипиды расщепляются до свободных жирных кислот, глицерола, фосфорной кислоты и аминоспирта или его аналога, например, аминокислоты серина, однако часть фосфолипидов расщепляется при участии фосфолипазы А2 только до лизофосфолипидов и в таком виде может поступать в стенку кишечника.
ФЛ А2 активируется частичным протеолизом с участием трипсина и гидролизует лецитин до лизолецитина. Лизолецитин является хорошим эмульгатором. ЛизоФЛ гидролизует часть лизолецитина до глицерофосфохолина. Остальные фосфолипиды не гидролизуются.
4. Гидролиз эфиров холестерина до холестерина и жирных кислот осуществляет холестеролэстераза, фермент поджелудочной железы и кишечного сока.
5. Мицеллообразование
Водонерастворимые продукты гидролиза (жирные кислоты с длинной цепью, 2-МГ, холестерол, лизолецитины, фосфолипиды) вместе с компонентами желчи (солями жёлчных кислот, ХС, ФЛ) образуют в просвете кишечника структуры, называемые смешанными мицеллами. Смешанные мицеллы построены таким образом, что гидрофобные части молекул обращены внутрь мицеллы (жирные кислоты, 2-МГ, 1-МГ), а гидрофильные (желчные кислоты, фосфолипиды, ХС) — наружу, поэтому мицеллы хорошо растворяются в водной фазе содержимого тонкой кишки. Стабильность мицелл обеспечивается в основном солями жёлчных кислот, а также моноглицеридами и лизофосфолипидами.
Регуляция переваривания
Пища стимулирует секрецию из клеток слизистой тонкой кишки в кровь холецистокинина (панкреозимин, пептидный гормон). Он вызывает выделение в просвет двенадцатиперстной кишки желчи из желчного пузыря и панкреатического сока из поджелудочной железы.
Кислый химус стимулирует секрецию из клеток слизистой тонкой кишки в кровь секретина (пептидный гормон). Секретин стимулирует секрецию бикарбоната (НСО3-) в сок поджелудочной железы.
Особенность переваривания липидов у детей
Секреторный аппарат кишечника к моменту рождения ребенка в целом сформирован, в кишечном соке находятся те же ферменты, что и у взрослых, но активность их низкая. Особенно напряженно идет процесс переваривания жиров из-за низкой активности липолитических ферментов. У детей, находящихся на грудном вскармливании, эмульгированные желчью липиды на 50% расщепляются под влиянием липазы материнского молока.
ВСАСЫВАНИЕ ПРОДУКТОВ ГИДРОЛИЗА
1. Водорастворимые продукты гидролиза липидов всасываются в тонкой кишке без участия мицелл. Холин и этаноламин всасываются в виде ЦДФ производных, фосфорная кислота – в виде Na+ и K+ солей, глицерол – в свободном виде.
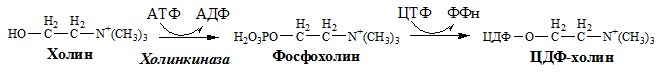
2. Жирные кислоты с короткой и средней цепью, всасываются без участия мицелл в основном в тонкой кишке, а часть уже в желудке.
3. Водонерастворимые продукты гидролиза липидов всасываются в тонкой кишке с участием мицелл. Мицеллы сближаются со щёточной каймой энтероцитов, и липидные компоненты мицелл (2-МГ, 1-МГ, жирные кислоты, холестерин, лизолецитин, фосфолипиды и т.д.) диффундируют через мембраны внутрь клеток.
Рециклирование компоненты желчи
Вместе с продуктами гидролиза всасываются компоненты желчи – соли жёлчных кислот, фосфолипиды, холестерин. Наиболее активно соли жёлчных кислот всасываются в подвздошной кишке. Жёлчные кислоты далее попадают через воротную вену в печень, из печени вновь секретируются в жёлчный пузырь и далее опять участвуют в эмульгировании липидов. Этот путь жёлчных кислот называют «энтерогепатическая циркуляция». Каждая молекула жёлчных кислот за сутки проходит 5— 8 циклов, и около 5% жёлчных кислот выделяется с фекалиями.
НАРУШЕНИЯ ПЕРЕВАРИВАНИЯ И ВСАСЫВАНИЯ ЛИПИДОВ. СТЕАТОРЕЯ
Нарушение переваривания липидов может быть при:
- 1) нарушение оттока жёлчи из жёлчного пузыря (желчекаменная болезнь, опухоль). Уменьшение секреции жёлчи вызывает нарушение эмульгирования липидов, что ведет к снижению гидролиза липидов пищеварительными ферментами;
- 2) нарушение секреции сока поджелудочной железы приводит к дефициту панкреатической липазы и снижает гидролиз липидов.
Нарушение переваривания липидов тормозит их всасывание, что приводит к увеличению количества липидов в фекалиях — возникает стеаторея (жирный стул). В норме в фекалиях липидов не более 5%. При стеаторее нарушается всасывание жирорастворимых витаминов (A, D, Е, К) и незаменимых жирных кислот (витамин F), поэтому развиваются гиповитаминозы жирорастворимых витаминов. Избыток липидов связывает вещества нелипидной природы (белки, углеводы, водорастворимые витамины), и препятствует их перевариванию и всасыванию. Возникают гиповитаминозы по водорастворимым витаминам, белковое и углеводное голодание. Непереваренные белки подвергаются гниению в толстой кишке.
ОБМЕН ЛИПИДОВ В ЭНТЕРОЦИТАХ
Липиды поступают в энтероциты как из просвета кишечника, так и из тканей. Большая часть липидов, поступивших в энтероцит, подвергается ресинтезу.
- 1. 1-МГ гидролизуется кишечной липазой до глицерина и жирной кислоты.
- 2. Короткоцепочечные жирные кислоты, ФЛ (кроме лецитина) и часть глицерина без изменений направляются из энтероцита в кровь.
- 3. Длинноцепочечные эндогенные и экзогенные жирные кислоты под действием ацил-КоА-синтетазы (тиокиназы) активируются, образуя Ацил
КоА:
RCOOH + HS-КоА + АТФ → Ацил
КоА + АМФ + ФФн
4. Моноацилглицероловый путь синтеза ТГ и ФЛ
При ресинтезе ТГ Ацил
КоА с участием ацилтрансферазы этерифицирует 2-МГ до ДГ, а затем до ТГ: 2-МГ + Ацил
КоА → 1,2-ДГ + HS-КоА,
1,2-ДГ + Ацил
КоА → ТГ + HS-КоА
При ресинтезе ФЛ на 1,2-ДГ переносится фосфохолин или фосфоэтаноламин с ЦДФ.
1,2-ДГ + ЦДФ-холин → лецитин + ЦМФ
1,2-ДГ + ЦДФ-этаноламин → кефалин + ЦМФ
В клетках слизистой оболочки тонкой кишки синтезируются в основном видоспецифичные ТГ. Однако при поступлении с пищей ТГ с необычными жирными кислотами, например бараньего жира, в адипоцитах появляются ТГ, содержащие кислоты, характерные для бараньего жира (насыщенные разветвлённые жирные кислоты).
5. Глицерофосфатный путь синтеза ТГ и ФЛ
глицерол + АТФ → глицеро-ф + АДФ Фермент: глицерокиназа
глицеро-ф + Ацил
КоА → лизофосфатид + HS-КоА Фермент: ацилтрансфераза
лизофосфатид + Ацил
КоА → фосфатид + HS-КоА Фермент: ацилтрансфераза
фосфатид + Н2О → 1,2-ДГ + Фн Фермент: фосфатидаза
1,2-ДГ + Ацил
КоА → ТГ + HS-КоА Фермент: ацилтрансфераза
1,2-ДГ + ЦДФ-холин → лецитин + ЦМФ Фермент: холинтрансфераза
1,2-ДГ + ЦДФ-этаноламин → кефалин + ЦМФ
6. При ресинтезе лецитина Ацил
КоА с участием ацилтрансферазы этерифицирует лизолецитин до лецитина: лизолецитин + Ацил
КоА → лецитин + HS-КоА
7. При ресинтез эфиров холестерина Ацил
КоА с участием ацилхолестеролацилтрансферазы (АХАТ) этерифицирует холестерин до эфира холестерина:
Ацил
КоА + ХС → ЭХС + HS-КоА
От активности АХАТ зависит скорость поступления экзогенного холестерола в организм.
ТРАНСПОРТ ЛИПИДОВ В ОРГАНИЗМЕ
Транспорт липидов в организме идет двумя путями:
1) жирные кислоты транспортируются в крови с помощью альбуминов;
2) ТГ, ФЛ, ХС, ЭХС и д.р. липиды транспортируются в крови в составе липопротеинов.
Обмен липопротеинов
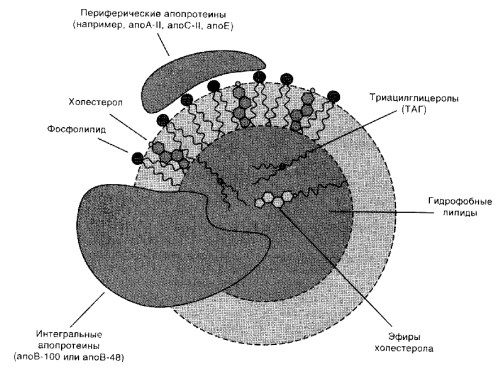
Липопротеины (ЛП) – это надмолекулярные комплексы сферической формы, состоящие из липидов, белков и углеводов. ЛП имеют гидрофильную оболочку и гидрофобное ядро. В гидрофильную оболочку входят белки и амфифильные липиды – ФЛ, ХС. В гидрофобное ядро входят гидрофобные липиды – ТГ, эфиры ХС и т.д. ЛП хорошо растворимы в воде.
В организме синтезируются несколько видов ЛП, они отличаются химическим составом, образуются в разных местах и осуществляют транспорт липидов в различных направлениях.
ЛП разделяют с помощью:
1) электрофореза, по заряду и размеру, на α-ЛП, β-ЛП, пре-β-ЛП и ХМ;
2) центрифугирования, по плотности, на ЛПВП, ЛПНП, ЛППП, ЛПОНП и ХМ.
Соотношение и количество ЛП в крови зависит от времени суток и от питания. В постабсорбтивный период и при голодании в крови присутствуют только ЛПНП и ЛПВП.
Транспорт к тканям экзогенных липидов пищи
Транспорт к тканям эндогенных липидов печени
Транспорт к тканям эндогенных липидов печени
Транспорт ХС в ткани
Удаление избытка ХС из тканей Донор а по А, С, Е
в крови из ЛПОНП
в крови из ЛППП
Апобелки
Белки, входящие в состав ЛП, называются апопротеины (апобелки, апо). К наиболее распространенным апопротеинам относят: апо А-I, А-II, В-48, В-100, С-I, С-II, С-III, D, Е. Апобелки могут быть периферическими (гидрофильные: А-II, С-II, Е) и интегральными (имеют гидрофобный участок: В-48, В-100). Периферические апо переходят между ЛП, а интегральные – нет.
Апопротеины выполняют несколько функций:
Активатор ЛХАТ, образование ЭХС
Активатор ЛХАТ, образование ЭХС
Структурная (синтез ЛП), рецепторная (фагоцитоз ЛП)
Структурная (синтез ЛП), рецепторная (фагоцитоз ЛП)
Активатор ЛХАТ, образование ЭХС
Активатор ЛПЛ, стимулирует гидролиз ТГ в ЛП
Ингибитор ЛПЛ, ингибирует гидролиз ТГ в ЛП
Перенос эфиров холестерина (БПЭХ)
Рецепторная, фагоцитоз ЛП
Ферменты транспорта липидов
Липопротеинлипаза (ЛПЛ) (КФ 3.1.1.34, ген LPL, около 40 дефектных аллелей) связана с гепарансульфатом, находящимся на поверхности эндотелиальных клеток капилляров кровеносных сосудов. Она гидролизует ТГ в составе ЛП до глицерина и 3 жирных кислот. При потере ТГ, ХМ превращаются в остаточные ХМ, а ЛПОНП повышают свою плотность до ЛППП и ЛПНП.
Апо С-II ЛП активирует ЛПЛ, а фосфолипиды ЛП участвуют в связывании ЛПЛ с поверхностью ЛП. Синтез ЛПЛ индуцируется инсулином. Апо С-III ингибирует ЛПЛ.
ЛПЛ синтезируется в клетках многих тканей: жировой, мышечной, в легких, селезёнке, клетках лактирующей молочной железы. Ее нет в печени. Изоферменты ЛПЛ разных тканей отличаются по значением Кm. В жировой ткани ЛПЛ имеет Кm в 10 раз больше, чем в миокарде, поэтому в жировая ткань поглощает жирные кислоты только при избытке ТГ в крови, а миокард – постоянно, даже при низкой концентрации ТГ в крови. Жирные кислоты в адипоцитах используются для синтеза ТГ, в миокарде как источник энергии.
Печёночная липаза находиться на поверхности гепатоцитов, она не действует на зрелые ХМ, а гидролизует ТГ в ЛППП.
Лецитин: холестерол-ацил-трансфераза (ЛХАТ) находиться в ЛПВП, она переносит ацил с лецитина на ХС с образование ЭХС и лизолецитина. Ее активируют апо А-I, А-II и С-I.
лецитин + ХС → лизолецитин + ЭХС
ЭХС погружается в ядро ЛПВП или переноситься с участием апо D на другие ЛП.
Рецепторы транспорта липидов
Рецептор ЛПНП — сложный белок, состоящий из 5 доменов и содержащий углеводную часть. Рецептор ЛПНП взаимодействует с белками ano B-100 и апо Е, хорошо связывает ЛПНП, хуже ЛППП, ЛПОНП, остаточные ХМ, содержащие эти апо. Клетки тканей содержат большое количество рецепторов ЛПНП на своей поверхности. Например, на одной клетке фибробласта имеется от 20 000 до 50 000 рецепторов.
Если количество холестерола, поступающего в клетку, превышает её потребность, то синтез рецепторов ЛПНП подавляется, что уменьшает поток холестерола из крови в клетки. При снижении концентрации свободного холестерола в клетке, наоборот, активируется синтез ГМГ-КоА-редуктазы и рецепторов ЛПНП. Стимулируют синтез рецепторов ЛПНП гормоны: инсулин и трийодтиронин (Т3), половые гормоны, а глюкокортикоиды – уменьшают.
Белок, сходным с рецептором ЛПНП на поверхности клеток многих органов (печени, мозга, плаценты) имеется другой тип рецептора, называемый «белком, сходным с рецептором ЛПНП». Этот рецептор взаимодействует с апо Е и захватывает ремнантные (остаточные) ХМ и ЛППП. Так как ремнантные частицы содержат ХС, этот тип рецепторов также обеспечивает поступление его в ткани.
Кроме поступления ХС в ткани путём эндоцитоза ЛП, некоторое количество ХС поступает в клетки путём диффузии из ЛПНП и других ЛП при их контакте с мембранами клеток.
В крови в норме концентрация:
ЛПНП
НОРМАЛЬНЫЕ ЗНАЧЕНИЯ
В соответствии c рекомендациями ASSMANN в настоящее время следующие значения холестерина ЛПВП могут быть предложены как основополагающие для оценки риска развития атеросклероза:
ОБМЕН ХИЛОМИКРОНОВ
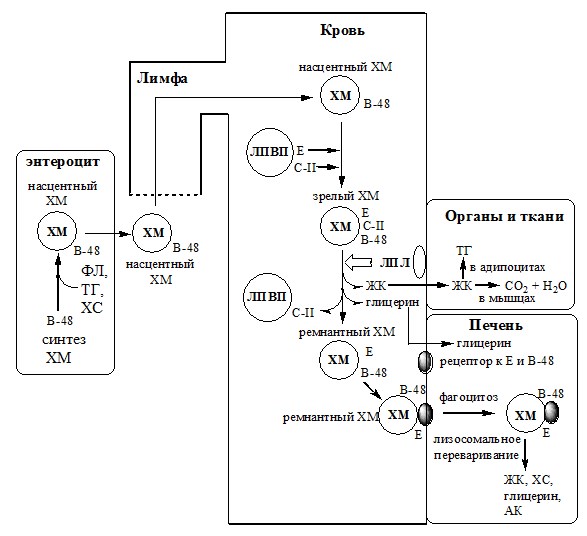
Липиды, ресинтезированные в энтероцитах, транспортируется тканям в составе ХМ.
- Образование ХМ начинается с синтеза апо В-48 на рибосомах. Апо В-48 и В-100 имеют общий ген. Если с гена копируется на мРНК только 48% информации, то с нее синтезируется апо В-48, если 100% – то с нее синтезируется апо В-100.
- С рибосом апо В-48 поступает в просвет ЭПР, где он гликозилируется. Затем в аппарате Гольджи апо В-48 окружается липидами и происходит формирование «незрелых», насцентных ХМ.
- Экзоцитозом насцентные ХМ выделяются в межклеточное пространство, поступают в лимфатические капилляры и по лимфатической системе, через главный грудной лимфатический проток попадают в кровь.
- В лимфе и крови с ЛПВП на насцентные ХМ переносятся апо Е и С-II, ХМ превращаются в «зрелые». ХМ имеют довольно большой размер, поэтому они придают плазме крови опалесцирующий, похожий на молоко, вид. Под действием ЛПЛ ТГ ХМ гидролизуются на жирные кислоты и глицерол. Основная масса жирных кислот проникает в ткань, а глицерол транспортируется с кровью в печень.
- Когда в ХМ количество ТГ снижается на 90%, они уменьшаются в размерах, а апо С-II переносится обратно на ЛПВП, «зрелые» ХМ превращаются в «остаточные» ремнантные ХМ. Ремнантные ХМ содержат в себе фосфолипиды, холестерол, жирорастворимые витамины и апо В-48 и Е.
- Ремнантные ХМ захватываются гепатоцитами, которые имеют рецепторы к апо В-48 и Е. Путём эндоцитоза остаточные ХМ попадают внутрь клеток и перевариваются в лизосомах. ХМ исчезают из крови в течение нескольких часов.
НАРУШЕНИЯ ОБМЕНА ХИЛОМИКРОНОВ
- 1. Абеталипопротеинемия (синдром Бассена-Корнцвейга)
При наследственном дефекте гена апо В — нарушается синтез апо В-100 в печени и апо В-48 в кишечнике. В результате в энтероцитах не формируются ХМ, а в печени — ЛПОНП. В клетках этих органов накапливаются капли ТГ, нарушается всасывание пищевых липидов.
В крови наблюдается полное отсутствие ХМ, ЛПОНП, ЛППП и ЛПНП, уменьшение количества ХС, ФЛ, высших полиненасыщенных жирных кислот, Е и А витаминов.
Уже на первом месяце жизни отмечаются плохой аппетит, обильные испражнения с признаками стеатореи, гипотрофия. Развивается акантоцитоз (акантоциты – эритроциты с множественными шиловидными выростами), анемия, аритмия. Вследствие демиелинизации аксонов возникает прогрессирующая атаксия, нистагм, пигментная дегенерация сетчатки и отставание в умственном развитии.
Атаксия — это нарушение координации движений при поражении лобных долей головного мозна, мозжечка, путей глубокой чувствительности в спинном и головном мозге. - 2. Семейная хиломикронемия ( гиперлипопротеинемии типа I) (менее 1% всех случаев гиперлипопротеинемии)
Генетические дефекты ЛПЛ или апо С-II вызывают нарушение катаболизма ХМ, приводящий к гиперхиломикронемии. Высокий уровень ХМ сохраняется в плазме крови спустя 12 ч после приема пищи. За счет избытка ХМ плазма крови по виду напоминает молоко, при отстаивании ХМ образуют на ее поверхности сливкообразный слой. Хиломикронемия сопровождается гипертриглицеролемией – уровень ТГ в крови может превышать 11,3 ммоль/л. Содержание ХС остается нормальным или повышается (гиперхолестеринемия). Коэффициент ХС/ТГ менее 0,2. Развитие атеросклероза не характерно.
Семейная хиломикронемия возникает у детей в возрасте до 10 лет. У больных происходит отложение ТГ в коже и сухожилиях в виде ксантом. Эруптивные ксантомы могут занимать большую часть поверхности кожи.
ТГ также откладываются в печени, селезенке, поджелудочной железе и других органах, что вызывает в этих органах сужение просвета сосудов, уменьшение кровотока, развитие тромбозов и ишемических некрозов. Развивается панкреатит, гепато- и спленомегалия.
У пациентов нарушается память, возникают абдоминальные боли, тошнота, рвота, желтушность кожи, липемия роговицы. Прогрессирование панкреатита часто бывает причиной смерти больных.
ОБМЕН β-ЛИПОПРОТЕИНОВ
В промежутках между приемами пищи и при голодании необходимые для тканей липиды синтезируются преимущественно в печени. Печень — основной орган, где идёт синтез жирных кислот, ХС, ФЛ из продуктов гликолиза. Скорость синтеза липидов в печени существенно зависит от состава пищи. Если в пище содержится более 10% липидов, то скорость синтеза липидов в печени резко снижается.
Транспорт липидов из печени осуществляется с участием ЛПОНП. Синтез ЛПОНП идет также как и ХМ. Сначала на рибосомах синтезируется апо В-100, который потом в аппарате Гольджи соединяется с липидами. Так как апо В-100 очень «длинный» белок (11536 АК), он покрывает поверхность всего ЛП.
После секреции ЛПОНП из печени в кровь, на них с ЛПВП переходят апо С-II и апо Е. Апо С-II активирует ЛПЛ, которая гидролизует ТГ ЛПОНП до жирных кислот и глицерина. Глицерол с кровью транспортируется в печень, а жирные кислоты – в ткань. Параллельно с потерей ТГ, ЛПОНП получают от ЛПВП ЭХС и ХС. В результате плотность ЛПОНП увеличивается, он превращается сначала в ЛППП, а затем в ЛПНП, при этом на ЛПВП возвращаются сначала апо С-II, а затем апо Е.
Содержание ЭХС и ХС в ЛППП достигает 45%; часть этих ЛП захватывается клетками печени через рецептор к ЛПНП (чувствителен к апо Е и апо В-100).
ЛПНП содержат до 55% ЭХС и ХС. ЛПНП являются основным поставщиком ХС в ткани. Из крови ЛПНП поступают в печень (до 75%) и другие ткани, которые имеют на своей поверхности рецепторы к ЛПНП.
НАРУШЕНИЯ ОБМЕНА β-ЛИПОПРОТЕИНОВ
- 1. Семейная β-липопротеинемия ( гиперлипопротеинемия типа IIа) (10% всех случаев гиперлипопротеинемии)
Возникает при дефекте апо В-100 (точечная мутация: арг замещен на глу), в результате чего уменьшается сродство ЛПНП к рецепторам В/Е, снижается катаболизм ЛПНП и увеличивается их концентрация в крови. β-липопротеинемия сопровождается гиперхолестеролемией, уровень ТГ в норме.
Семейный дефект апопротеида В-100 встречается с несколько меньшей частотой, чем семейная гиперхолестеринемия, и отличается от нее отсутствием сухожильных ксантом и более низким уровнем ХС. - 2. Семейная гиперхолестеролемия (гиперлипопротеинемия типа IIа и IIв)
Наследственный дефект рецептора ЛПНП (к апо В/Е) (или белка апоВ-100) приводит к развитию распространённого наследственного заболевания — семейной гиперхолестеролемии.
При дефекте рецептора ЛПНП наблюдается триглицеролемия, при дефекте белка апоВ-100 – нет.
У гетерозигот (1:400) количество рецепторов ЛПНП на поверхности клеток снижено вдвое, а концентрация ХС в плазме примерно вдвое выше нормы (9-12 ммоль/л). ЛПНП фагоцитируются макрофагами. Нагруженные избытком ХС и других липидов, макрофаги откладываются в коже, сухожилиях и образуют ксантомы. Гиперхолестеролемия приводит к выраженному атеросклерозу, развитию ИБС и ранней смерти в результате инфаркта миокарда или инсульта.
У гомозигот (1:1000000) нет рецепторов к ЛПНП, концентрация ХС и ЛПНП в крови уже в раннем детском возрасте превышает норму в 5-6 раз (20-40 ммоль/л). Для пациентов характерно наличие не только ксантоматоза сухожилий, но и эруптивных ксантом на ягодицах, коленях, локтях, слизистой оболочке полости рта.
Избыток в крови ХС и ЛПНП способствует быстрому развитию атеросклероза и ИБС. Такие дети без экстренных мер лечения погибают в возрасте 5—6 лет.
ОБМЕН ЛПВП
ЛПВП выполняют 2 основные функции: они поставляют апо другим ЛП в крови и участвуют в так называемом «обратном транспорте ХС». ЛПВП синтезируются в печени и в небольшом количестве в тонком кишечнике в виде насцентных ЛПВП. Они имеют дисковидную форму, небольшой размер и содержат высокий процент белков и фосфолипидов. В печени в ЛПВП включаются апопротеины А, Е, С-II, ЛХАТ. В крови апо С-II и апо Е переносятся с ЛПВП на ХМ и ЛПОНП. насцентные ЛПВП практически не содержат ХС и ТГ и в крови обогащаются ХС, получая его из других ЛП и мембран клеток.
Для переноса ХС в ЛПВП существует сложный механизм. На поверхности ЛПВП находится фермент ЛХАТ — лецитин: холестерол-ацилтрансфераза. Этот фермент превращает ХС в ЭХС. Реакция активируется апо A-I, входящим в состав ЛПВП.
ЭХС перемещается внутрь ЛПВП. Таким образом, ЛПВП обогащаются ЭХС. ЛПВП увеличиваются в размерах, из дисковидных небольших частиц превращаются в частицы сферической формы, которые называют ЛПВП3, или «зрелые ЛПВП». ЛПВП3 частично обменивают ЭХС на ТГ, содержащиеся в ЛПОНП, ЛППП и ХМ. В этом переносе участвует «белок, переносящий эфиры холестерина» – апо D. Таким образом, часть ЭХС переносится на ЛПОНП, ЛППП, а ЛПВП3 за счёт накопления ТГ увеличиваются в размерах и превращаются в ЛПВП2.
Часть ЛПВП захватывается клетками печени, взаимодействуя со специфическими для ЛПВП рецепторами к апо А-1. На поверхности клеток печени ФЛ и ТГ ЛППП, ЛПВП2 гидролизуются печёночной липазой, что дестабилизирует структуру поверхности ЛП и способствует диффузии ХС в гепатоциты. ЛПВП2 в результате этого опять превращаются в ЛПВП3 и возвращаются в кровоток.
НАРУШЕНИЯ ОБМЕНА ЛПВП
Болезнь Тэнжи
Болеют аборигены острова Тэнжи. Наследственный дефект апо А, не синтезируются ЛПВП. Нарушается транспорт излишков ХС из тканей в печень. В крови низкий уровень ХС, ФЛ, много ТГ. Макрофаги фагоцитируют в тканях излишки ХС с образованием ксантом. Накопление ХС в печени, селезенке и других лимфоидных органах вызывает гепатоспленомегалию и лимфаденопатию. Может развиваться катаракта, полинейропатия и ренит. Миндалины из-за отложений ХС окрашены в оранжево-желтый цвет.

Белая и бурая жировая ткань. Липолиз и липогенез.
Жировая ткань – белая и бурая: особенности химического состава, обмена белков, жиров, углеводов, энергетического метаболизма.
Жировая ткань – это разновидность соединительной ткани, составляет 15-20% веса мужчин и 20-25% веса женщин. Как и любая ткань, она состоит из клеток (адипоциты, макрофаги, тучные клетки и т.д.) и межклеточного вещества. Межклеточное вещество в жировой ткани представлено небольшим количеством волокон (коллагена и эластина) и основного вещества (гликозаминогликаны, протеоликаны).
Жировая ткань делиться на белую и бурую:
📺 Видео
Гидролиз солей. 9 класс.Скачать

РЕАКЦИИ ИОННОГО ОБМЕНА, ИОННОЕ УРАВНЕНИЕ - Урок Химия 9 класс / Подготовка к ЕГЭ по ХимииСкачать

Ступенчатый гидролиз солей по аниону. Решаем примеры.Скачать

Химия 9 класс (Урок№8 - Гидролиз солей.)Скачать

Гидролиз солей. Теория для задания 23 ЕГЭ по химии.Скачать

11 класс. Гидролиз солей.Скачать

Всё про ГИДРОЛИЗ для ЕГЭ ПО ХИМИИ. с ЭКСПЕРИМЕНТАМИ!Скачать

Гидролиз солей.ПримерыСкачать

Химические уравнения // Как Составлять Уравнения Реакций // Химия 9 классСкачать

ЭЛЕКТРОЛИТИЧЕСКАЯ ДИССОЦИАЦИЯ ХИМИЯ 8 класс // Подготовка к ЕГЭ по Химии - INTENSIVСкачать

Вычисление рН в растворах солейСкачать

Гидролиз солей | ХимияСкачать

Степень гидролиза и как её рассчитатьСкачать

Гидролиз солейСкачать

Электролиз. 10 класс.Скачать

Химия 11 класс (Урок№7 - Гидролиз органических и неорганических соединений.)Скачать

8 класс. ОВР. Окислительно-восстановительные реакции.Скачать

Химия 8 класс: Гидролиз солейСкачать
