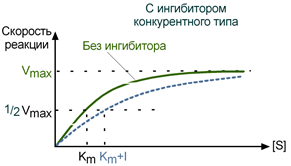
При таком виде ингибирования ингибитор конкурирует с субстратом за активный центр (за контактный участок), что приводит к уменьшению связывания субстрата с ферментом и снижению катализа. Используя терминологию кинетики Михаэлиса-Ментен можно сказать, что конкурентный ингибитор уменьшает сродство фермента к субстрату, повышая константу Михаэлиса (Km), максимальная скорость реакции (Vmax) остается при этом неизменнной.
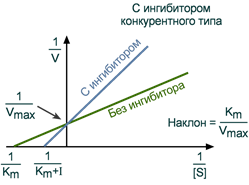
Зависимость в координатах Лайнуивера-Берка имеет вид пучка прямых, пересекающихся на оси ординат. Конкурентный ингибитор увеличивает Кm и не изменяет Vmax.
График конкурентного ингибирования в координатах Михаэлиса-Ментен
График конкурентного ингибирования в координатах Лайнуивера-Берка
Кинетика неконкурентного ингибирования
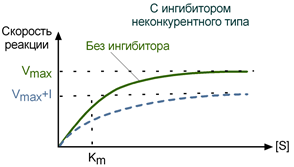
Особенностью неконкурентного ингибитора является его способность связываться с ферментом не в активном центре, и изменение концентрации субстрата никак не влияет на это связывание. В то же время неконкурентный ингибитор не мешает связыванию субстрата с активным центром. В результате формируется тройной комплекс фермент-субстрат-ингибитор (E-S-I), в котором фермент уже не способен изменить свою конформацию и обеспечить проведение реакции. Количество «работоспособных» комплексов E-S при этом снижается.
Максимальная скорость реакции (Vmax) при неконкурентном ингибировании снижается, константа Михаэлиса (Km) не изменяется, т.е. добавление дополнительного субстрата не может повлиять на состояние активного центра и работу фермента.
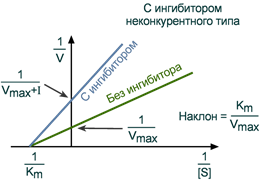
Зависимость в координатах Лайнуивера-Берка имеет вид пучка прямых, пересекающихся на оси абсцисс. Неконкурентный ингибитор не изменяет Кm и снижает Vmax.
График неконкурентного ингибирования в координатах Михаэлиса-Ментен
График неконкурентного ингибирования в координатах Лайнуивера-Берка
Кинетика бесконкурентного ингибирования
Бесконкурентный ингибитор способен связываться только с уже образовавшимся комплексом E-S, но не со свободным ферментом. Связывание происходит либо на молекуле субстрата, либо с ферментом, уже начавшем катализ и изменившим свою конформацию. Образовавшийся тройной комплекс E-S-I также непродуктивен.
Максимальная скорость реакции (Vmax) и константа Михаэлиса (Km) при бесконкурентном ингибировании снижаются. Добавление дополнительного субстрата не может повлиять на состояние активного центра и работу фермента.
Графический анализ конкурентного ингибирования по уравнению Лайнуивера-Берка
КОНКУРЕНТНОЕ ИНГИБИРОВАНИЕ- когда за активный центр фермента вместе с субстратом конкурирует ИНГИБИТОР:
Е + I = EI Ki = [E][I] / [EI], чем меньше Ki, тем сильнее ингибитор.
скор р-ции.
Зависимость в координатах Лайнуивера-Берка имеет вид пучка прямых, пересекающихся на оси ординат. КОНКУРЕНТНЫЙ ИНГИБИТОР увеличивает Кm и не изменяет V max.
Ингибирование происходит из-за образование фермент-субстратного комплекса и уменьшении концентрации фермента в системе
-30 – без ингибирования
-20 – с уменьшением концентрации ингибирования
-5 — с увеличением концентрации ингибирования
Графический анализ неконкурентного ингибирования по уравнению Лайнуивера-Берка
НЕКОНКУРЕНТНОЕ ИНГИБИРОВАНИЕ ПРОЯВЛЯЕТСЯ ПРИ СВЯЗЫВАНИИ ИНГИБИТОРА С ФЕРМЕНТОМ ВНЕ АКТИВНОГО ЦЕНТРА, но при этом меняется структура активного центра и связь с субстратом становится невозможной.
E + I —> EI, EI + S —-> (невозможно)
скор р-ции
Зависимость в координатах Лайнуивера-Берка имеет вид пучка прямых, пересекающихся на оси абсцисс. ингибитор не изменяет Кm и снижает V max.
Нет разницы сколько субстрата в системе
53 Определение и физический смысл константы ингибирования (Кi) ферментативных реакций
1) КОНКУРЕНТНЫЙ ИНГИБИТОР увеличивает Кm и не изменяет V max.
2) НЕКОНКУРЕНТНЫЙ ингибитор не изменяет Кm и снижает V max.
3) БЕСКОНКУРЕНТНЫЙ ингибитор в одинаковой степени снижает Кm и V max:
КОНКУР ИНГИБИРОВАНИЕ: (α → ∞, β не имеет определенного смысла).
Зависимость в координатах Лайнуивера-Берка имеет вид пучка прямых, пересекающихся на оси ординат.
Константу конкурентного ингибирования Кi можно определить, зная величину
если отложить экспериментальные данные в координатах (КМ(каж), [I]).
неконкурентное ингибирование(α → 1, β =0)
(6.5)
Зависимость в координатах Лайнуивера-Берка имеет вид пучка прямых, пересекающихся на оси абсцисс.
Константу неконкурентного ингибирования Кi можно определить, зная величину
(6.6)
если представить экспериментальные данные в координатах (1/kkat, [I])
Бесконкурентное ингибирование(α=β
Механизмы взаимодействия эффекторов с ферментами.
Лекция №7
Изучение ферментативных реакций
Определение Vmax и Km. Линейные анаморфозы уравнения Михаэлиса-Ментен. Уравнения Лайнуивера-Бэрка и Иди-Хофсти. Механизмы взаимодействия эффекторов с ферментами. Ингибиторы и активаторы ферментов. Типы ингибирования. Кинетика ингибирования
Определение Vmax и Km. Линейные анаморфозы уравнения Михаэлиса-Ментен.
Уравнения Лайнуивера-Бэрка и Иди-Хофсти.
В принципе Vmax и Kmможно определить по графику уравнения Михаэлиса-Ментен, но так как Vmaxявляется ассимптотой, то затруднительно получить надежное значение Vmax и Kmпутем экстраполяции. И так главная задача состоит в том, чтобы с максимальной точностью определить параметры Vmax и Km. С этой целью используются два уравнения: уравнение Лайнуивера-Бэрка и Иди-Хофсти. Они представляют собой линейные анаморфозы уравнения Михаэлиса-Ментен. Г. Лайнуивер и Д. Бэрк преобразовали уравнение Михаэлиса-Ментен по методу двойных обратных величин исходя из того принципа, что если существует равенство между двумя какими-либо величинами, то и обратные величины также будут равны. В частности, если
после преобразования получаем уравнение:
которое получило название уравнения Лайнуивера–Бэрка. Это уравнение прямой линии: у = ах + b. Если теперь в соответствии с этим уравнением построить график в координатах 1/v (y) от l/[S] (x), то получим прямую линию (см.рис.), тангенс угла наклона который будет равен величине Km/Vmax; отрезок, отсекаемый прямой от оси ординат, представляет собой l/Vmax(обратная величина максимальной скорости). Если продолжить прямую линию за ось ординат, тогда на абсциссе отсекается отрезок, соответствующий обратной величине константы Михаэлиса – 1/Кm . Таким образом, величину Кm можно вычислить из данных наклона прямой и длины отрезка, отсекаемого от оси ординат, или из длины отрезка, отсекаемого от оси абсцисс в области отрицательных значений.
График Лайнуивера-Бэрка.
Следует подчеркнуть, что данный метод нашел широкое применение в современной энзимологии.
Уравнение Иди-Хофсти получается в результате следующих преобразований:
Поделим левую и правую части уравнения Михаэлиса-Ментен на [S] и получим:
после преобразований получаем:
т.е уравнение прямой линии типа У = — аХ + b: где У = v/[S], Х = v, а = — 1/Km, b = Vmax/Km
Это уравнение целесообразно преобразовать к еще более удобному виду умножив обе
части уравнения на Km, тогда получаем следующее уравнение, которое также называется уравнением Иди-Хофсти:
График этого уравнения представлен ниже.
Основные константы ферментативной реакции легко определяются из графиков Лайнуивера-Берка и Иди-Хофсти.
Размерность максимальной скорости –нкат.
Уравнения Лайнуивера-Берка и Иди-Хофсти удобны не только для точного определения Vmax и Km, но и для определения характера влияния различных агентов на скорость ферментативной реакции, т.е. определения влияния различных веществ на ферменты.
Действительно, если вещество каким-либо образом взаимодействует с ферментом, то это находит отражение в графиках кинетических зависимостей. Причем по характеру, наблюдающихся изменений кинетических зависимостей можно судить о характере взаимодействия фермента с веществом. В любом случае при этом изменяется ферментативная активность.
Механизмы взаимодействия эффекторов с ферментами.
Вещества, которые оказывают влияние на активность ферментов, называют эффекторами. Эффекторы делятся на ингибиторы и активаторы.
Активаторы ферментов
Активаторами ферментовназывают соединения, которые обуславливают или увеличивают активность фермента, но которые в отличие от кофермента сами по себе не принимают участие в реакции. По механизму активации они делятся на три группы:
1. взаимодействующие с активным центром фермента
2. взаимодействующие с аллостерическими участками фермента
3. взаимодействующие с субстратом
Взаимодействующие с активным центром ферментов активаторы называют обычно кофакторами ферментов– это обычно ионы металлов. В активных центрах многих ферментов содержаться такие ионы как Zn +2 , Co +2 , Cu +2 , Mn +2 и др. без которых фермент не работает. Кроме того, для стабилизации специфической конформации фермента, обуславливающей его максимальную каталитическую активность необходимы также многие одновалентные ионы Na + , K + , NH4 + .
Вторая группа активаторов, взаимодействующих с аллостерическими участками ферментов, называетсяаллостерическими активаторами. Это вещества, которые взаимодействуют с дополнительными, регуляторными центрами, несовпадающими с активным центром, вызывая изменение конформации в районе активного центра, так что он собственно формируется, и таким образом активируют фермент.
Известен и третий тип активаторов, взаимодействующих с субстратом. Например, для большинства реакций протекающих с участием АДФ и АТФ и других ди- тринуклеозид фосфатов необходимо присутствие в реакционной смеси стехеометрических количеств ионов двухвалентных металлов (обычно Mg +2 ). Исследование таких реакций показало, что механизм такой активации состоит в образовании комплекса катиона с субстратом до его взаимодействия с ферментом. Таким образом, истинным реагирующим веществом является комплекс субстрата с ионом металла, а не свободный субстрат.
Изучение кинетики ферментативных реакций позволяет выяснить влияние эффекторов на скорость ферментативной реакции и на основании этого судить о характере взаимодействия эффектора и фермента. На рисунке 2 представлены данные по влиянию 2-х активаторов на скорость ферментативной реакции, представленные на зависимости Лайнуивера-Берка.
Рис.2. Изменение характера кинетической кривой при активировании фермента при постоянной концентрации активатора и изменении концентрации субстрата.
(по графику Лайнуивера-Берка)
На графике Иди-Хофсти эти же зависимости будут выглядеть следующим образом: (см рис слева)
В том случае когда имеет место аллостерическое активирование кинетика не подчиняется уравнению Михаэлиса-Ментен и графики Лайнуивера-Берка и Иди-Хофсти применять нельзя.
В этом случае график зависимости начальной скорости реакции от концентрации субстрата имеет не гиперболическую форму, а сигмоидный характер (см. рис.3). Это означает, что связывание одной молекулы субстрата в одном каталитическом центре повышает связывание субстрата с другим центром, т.е. имеет место кооперативное взаимодействие, как, например, в случае присоединения кислорода к 4 субъединицам гемоглобина. Сигмоидный характер имеют и зависимости в том случае, если активатор воздействует на аллостерический участок фермента таким образом, что это собственно и приводит к формированию активного центра.
Рис. 3. Сигмоидная кинетическая
Кривая.
Изучение механизмов активирования ферментов привело к развитию представлений о характере взаимодействия субстратов с ферментами. Современные представления об этом сформулировал, главным образом, биохимик Кошланд.Суть его теориииндуцированного взаимодействия субстрата и фермента заключается в том, что активный центр фермента формируется в момент взаимодействия субстрата с ферментом, а не присутствует изначально в молекуле фермента, как полагали ранее.
Для оценки концентрации субстрата, при которой скорость реакции составляет половину максимальной (для определения Кm), в условиях сигмоидного характера кинетической кривой обычно применяют уравнение Хилла:
где К’ – константа Михаэлиса; n – число субстрат-связывающих центров.
Ингибиторы ферментов
Наибольший интерес для фармакологии и фармацевтической химии представляют ингибиторы ферментов.
Многие лекарственные вещества являются ингибиторами ферментов. Например, ингибиторы амилаз успешно применяются для лечения заболеваний, связанных с повышенной активностью этих ферментов – диабета, ожирения, кариеса. Используемые в военном деле нервно-паралитические газы – фосфорорганические соединения представляют собой специфические ингибиторы фермента холинэстеразы.
В научных исследованиях специфические ингибиторы используются для изучения механизма действия ферментов, строения их активного центра. Например, многие из промежуточных продуктов гликолиза и дрожжевого брожения были открыты благодаря использованию ингибиторов, блокирующих последовательные стадии процесса. В результате такого блокирования соответствующие промежуточные продукты накапливались в количествах, достаточных для их выделения и идентификации.
По характеру действия ингибиторы делятся на обратимые, необратимые и аллостерические. Удаление обратимых ингибиторов из системы (диализом, гельфильтрацией и др.) восстанавливает каталитическую активность фермента.
Необратимые ингибиторы необратимо модифицируют фермент, вследствие чего он полностью и навсегда выводится из строя – полностью теряет свою каталитическую активность.
Аллостерические ингибиторы воздействуют на аллостерические участки фермента, что приводит к изменению активного центра фермента и снижению его ферментативной активности.
Механизмы действия ингибиторов на ферменты определяют путем сравнения кинетики реакции в присутствии ингибитора и без него.
Обратимые ингибиторы (конкурентное взаимодействие с ферментом)
Рассмотрим вначале пример моносубстратной реакции в которой скорость реакции снижается по мере накопления продукта реакции. Этот эффект носит название ингибирование продуктом реакции.Такое ингибирование указывает на то, что процесс образования продукта из субстрата является обратимым, что справедливо для большинства ферментативных реакций. Маловероятно, что фермент, способный образовать с субстратом комплекс, подвергающийся обратимому превращению в продукт, не был в то же время способен образовать обратимый комплекс с продуктом. Механизм, в котором учитывается это обстоятельство можно записать следующим образом:
Е + S ↔ ES ↔ EP ↔ P + E
Из этой модели видно, что увеличение концентрации Р приводит к уменьшению концентрации свободного фермента, вследствие образования комплекса ЕР. Такое ингибирование называется конкурентным ингибированием – за связывание с ферментом конкурирует и субстрат и продукт реакции. На рисунке 4 представлено влияние этого эффекта на графиках Лейнуивера-Берка и Иди-Хофсти.
Примером конкурентного ингибирования продуктом реакции является ингибированияе спиртового брожения накапливающимся спиртом. Накопление этилового спрта в ходе брожения замедляет скорость реакции гидрирования ацетальдегида под действием фермента алкоголь дегидрогеназы и, наконец в системе устанавливается равновесие и накопление спирта в системе прекращается при достижении концентрации этанола в системе около15% об.
Конкурентное ингибирование может быть не только под действием продукта реакции но и при действии постороннего ингибитора, добавленного в реакционную смесь, например лекарственного вещества. В этом случае изучение кинетики реакции при определенной закрепленной концентрации ингибитора дает туже картину (Vmax = const, Km возрастает).
Таким образом суть конкурентного ингибирования заключается в конкурировании за активный центр фермента. По строению конкурентные ингибиторы могут быть аналогами субстрата целевого фермента или аналогами переходного состояния. Например, сульфаниламиды являются аналогами пара-аминобензойной кислоты, которая является субстратом фермента фолат-синтетазы. Сульфамиды обратимо связываются с активным центром фермента и таким образом обратимо ингибируют синтез фолиевой кислоты, необходимой бактериям для синтеза белка. Поэтому сульфаниламиды оказывают ингибирующее действие на рост бактерий, но не убивают их (бактериостатическое действие). Так 1 г пара-аминобензойной кислоты нивелирует действие 270 г норсульфазола.
Необратимые ингибиторы (неконкурентное взаимодействие с ферментом)
Аналоги субстрата могут и необратимо связываться с активным центром фермента (их называют суицидными субстратами) полностью необратимо выводя фермент из реакции. Такие аналоги субстрата оказывают неконкурентное необратимое ингибирование. На кинетических кривых это отражается следующим образом (см. рис.5)
Необратимую инактивацию фермента вызывают соединения (найденные в живой природе или полученные путем синтеза), которые вступают в химическую реакцию с образованием ковалентных связей с функциональными группами активного центра фермента. Такие соединения, специфически реагирующие с определенными группами в молекулах ферментов (групп-специфические реагенты), используют для идентификации функциональных групп активного центра (метод химической модификации).
С этой целью широко используются соединения, блокирующие SH-группы (иодацетамид, n-хлормеркурбензоат натрия и др.), окисляющие остатки триптофана в кислой среде (N-бромсукцинимид), ацетилирующие остатки тирозина (N-ацетилимидазол), связывающие металлы (азид натрия) и т.д.
К ним относятся и многие яды (в т. ч. пестициды), β-лактамные антибиотики (пенициллины, цефалоспорины и др.). Избирательность действия ингибиторов объясняется тем, что они реагируют со строго определенной химической группировкой в активном центре фермента. Например, соли синильной кислоты, окись углерода, азид натрия избирательно реагируют с некоторыми металлами, главным образом с железом и медью, и поэтому ингибируют многие железо- и медьсодержащие ферменты, к числу которых относятся так называемые дыхательные ферменты — ферменты тканевого дыхания. Соли ртути, препараты мышьяка, йодацетамид и некоторые другие вещества специфически реагируют с сульфгидрильными группами, угнетая ферменты, в состав активного центра которых входят SH-группы. Многие инсектициды, а также ядовитые вещества нервно-паралитического действия избирательно взаимодействуют с ОН-группой аминокислоты серина и благодаря этому резко тормозят активность холинэстеразы и некоторых других эстераз.
 скор р-ции.
скор р-ции.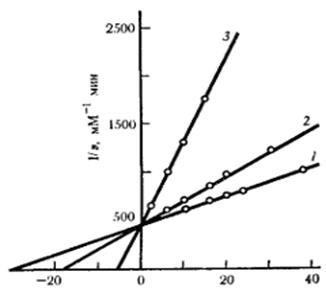
 скор р-ции
скор р-ции



 (6.6)
(6.6)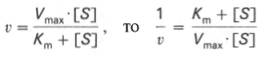




 На графике Иди-Хофсти эти же зависимости будут выглядеть следующим образом: (см рис слева)
На графике Иди-Хофсти эти же зависимости будут выглядеть следующим образом: (см рис слева) В том случае когда имеет место аллостерическое активирование кинетика не подчиняется уравнению Михаэлиса-Ментен и графики Лайнуивера-Берка и Иди-Хофсти применять нельзя.
В том случае когда имеет место аллостерическое активирование кинетика не подчиняется уравнению Михаэлиса-Ментен и графики Лайнуивера-Берка и Иди-Хофсти применять нельзя.

