Цикл Кальвина
Восстановительный пентозофосфатный цикл, или цикл Кальвина, — серия биохимических реакций, осуществляемая при фотосинтезерастениями (в стромехлоропластов), цианобактериями, прохлорофитами и пурпурными бактериями, а также многими бактериями-хемосинтетиками, является наиболее распространённым из механизмов автотрофной фиксации CO2.
Цикл Кальвина назван в честь американского биохимика Мелвина Кальвина (1911—1997). Часто используются альтернативные названия, указывающие на роль коллег Кальвина в открытии данного биохимического пути (например: цикл Кальвина-Бенсона или цикл Кальвина-Бенсона-Бассама).
Стадии
В цикл вовлекаются АТФ и НАДФ·Н, образованные в ЭТЦ фотосинтеза, углекислый газ и вода; основным продуктом является глицеральдегид-3-фосфат.
Поскольку АТФ и НАДФ·Н могут образовываться в разных метаболических путях, цикл не следует рассматривать строго привязанным к световой фазе фотосинтеза.
Общий баланс реакций цикла можно представить уравнением:
3 CO2 + 6 НАДФ·Н + 5 H2O + 9 АТФ → C3H7O3-PO3 + 3 H+ + 6 НАДФ+ + 9 АДФ + 8 Фн + 3 H2O
Две молекулы глицеральдегид-3-фосфата используются для синтеза глюкозы.
Цикл состоит из трёх стадий: на первой под действием ферментарибулозобисфосфат-карбоксилаза/оксигеназа происходит присоединение CO2 к рибулозо-1,5-дифосфату и расщепление полученной гексозы на две молекулы 3-фосфоглицериновой кислоты (3-ФГК). На второй 3-ФГК восстанавливается до глицеральдегид-3-фосфата (фосфоглицеральдегида, ФГА), часть молекул которого выходит из цикла для синтеза глюкозы, а другая часть используется в третьей стадии для регенерации рибулозо-1,5-дифосфата.
Карбоксилирование
Карбоксилированиерибулозо-1,5-бисфосфата (5-углеродное соединение) осуществляется РиБисКО в несколько стадий.
На первой кетонная группа рибулозы восстанавливается до спиртовой, между 2 и 3 атомами углерода устанавливается двойная связь. Полученное соединение нестабильно и именно оно карбоксилируется с образованием 2-карбокси-3-кето-D-арабитол-1,5-бисфосфата. Его структурный аналог 2-карбокси-D-арабитол-1,5-бисфосфат ингибирует весь процесс.
Новое, уже 6-углеродное соединение, также нестабильно и распадается на две молекулы 3-фосфоглицериновой кислоты (3-фосфоглицерат, 3-ФГК).
Восстановление
Восстановление 3-фосфоглицериновой кислоты (3-ФГК) происходит в две реакции.
Сначала каждая 3-ФГК с помощью 3-фосфоглицераткиназы и с затратой одной АТФ фосфорилируется, образуя 1,3-бисфосфоглицериновую кислоту (1,3-бисфосфоглицерат).
Затем под действием глицеральдегид-1,3-фосфатдегидрогеназы бисфосфоглицериновая кислота восстанавливается НАД(Ф)·H (у растений и цианобактерий; у пурпурных и зелёных бактерий восстановителем является НАД·H) параллельно с отщеплением одного остатка фосфорной кислоты.
Образуется глицеральдегид-3-фосфат (фосфоглицеральдегид, ФГА, триозофосфат). Обе реакции обратимы.
Регенерация
На последней стадии 5 молекул глицеральдегид-3-фосфатов превращаются в три молекулы рибулозо-1,5-бисфосфата.
Вначале под действием трифосфат-изомеразы глицеральдегид-3-фосфат изомеризуется в дигидроксиацетон-фосфат. Фруктозабисфосфат-альдолаза объединяет их во фруктозо-6-фосфат с отщеплением остатка фосфорной кислоты.
Затем следует ряд реакций перестройки углеродных скелетов и образуется рибулозо-5-фосфат. Он фосфорилируется фосфорибулокиназой и рибулозо-1,5-бисфосфат регенерирует.[источник не указан 2801 день]
Открытие
С 1940-х гг. Мелвин Кальвин работал над проблемой фотосинтеза; к 1957 с помощью CO2, меченного по углероду, выяснил химизм усвоения растениями CO2 (восстановительный карбоновый цикл Кальвина) при фотосинтезе. Нобелевская премия по химии (1961).
 Схема цикла.
Схема цикла.
Чёрные кружки — атомы углерода, красные — кислорода, фиолетовые — фосфора, маленькие чёрные окружности — атомы водорода
За световой фазой следует темновая фаза фотосинтеза, во время которой происходит синтез моносахаридов (глюкозы) из углекислого газа с затратой энергии АТФ и восстановительных эквивалентов (НАДФН). Синтез глюкозы является результатом целого ряда последовательных ферментативных реакций, которые назвали циклом Кальвина.
Как было сказано ранее в разделе «Кислородный этап энергетического обмена», в цикле Кребса в митохондриях от молекул органических кислот отрываются молекулы углекислого газа (CO2), промежуточные продукты цикла последовательно окисляются, отрываемые от них атомы водорода присоединяются к НАД+ (т.е. образуется НАДН). В цикле Кальвина происходит все наоборот, к молекулам субстрата присоединяется молекулы углекислого газа (СО2), и они восстанавливаются за счет НАДФН (т.е образуется НАДФ+).
Началом синтеза глюкозы является присоединение молекулы углекислого газа к молекуле пятиуглеродного сахара – рибулозо-1,5-бисфосфата.
При этом образуется шестиуглеродная молекула, которая сразу же распадается на две молекулы трехуглеродной фосфоглицериновой кислоты, которая восстанавливается до трехуглеродных сахаров с затратой АТФ и НАДФН. В результате их дальнейших перестроек и конденсаций образуются рибулозомонофосфат и глюкоза — конечный продукт фотосинтеза. Рибулозомонофосфат фосфорилируется АТФ до рибулозобисфостата, который вновь вступает в цикл Кальвина.
На образование одной молекулы глюкозызатрачивается 18 молекул АТФ и 12 молекул НАДФН, накопленных в процессе световой фазы фотосинтеза.
Какие основные процессы происходят в темновую фазу фотосинтеза?
Следовательно, для темновой фазы фотосинтеза можно представить следующее общее уравнение:
6СО2 + 12НАДФН + 12Н+ + 18АТФ —> С6Н12О6 + 6Н2О + 12НАДФ+ + 18АДФ + 18Фн
Даже если учесть частичные потери энергии на различных стадиях темновой фазы, общий КПД фотосинтеза остается очень высоким и составляет приблизительно 60%.
У некоторых растений (например, сахарного тростника или кукурузы) процесс фотосинтеза идет вначале не через трехуглеродные, а через четырехуглеродные соединения.
Эти растения называются С4-растениями. В отличие от С3-растений им характерен быстрый рост и высокая эффективность фотосинтеза, который протекает даже при очень низких концентрациях углекислого газа. В этом случае углекислый газ присоединяется не к рибулозобисфосфату, а к одному из промежуточных продуктов гликолиза – фосфоенолпирувату.
В результате образуются четырехуглеродные яблочная или аспарагиновая кислоты, которые диффундируют в клетки обкладки сосудистых пучков, где от них отщепляется СО2, вступая в цикл Кальвина.
В этих клетках слабо выражено фотодыхание, связанное с окислением рибулозобисфосфата кислородом, поэтому энергозатраты на фотосинтез резко снижаются (на 50%).
В последние годы благодаря необычайно высокой биологической продуктивности С4-растения привлекают внимание ученых как потенциальный источник органического сырья.
Темновая фаза фотосинтеза
Темновая фаза фотосинтеза – это комплекс ферментативных реакций, во время которой происходит восстановление поглощенного углекислого газа за счет продуктов световой фазы (АТФ и НАДФН). Различают несколько циклов восстановления СО2.
Этот способ ассимиляции СО2 является основным и присущ всем растениям. Он был расшифрован американскими учеными во главе с М. Кальвином. В 1961 году М. Кальвину за установление последовательности реакций в этом цикле и была присуждена Нобелевская премия.
Этот цикл начинается с присоединения СО2 к акцептору – пятиуглеродному сахару рибулозо-1,5-дифосфату (РДФ).
Присоединение СО2 к тому или ионному веществу называется карбоксилированием, а фермент катализирующий такую реакцию – карбоксилазой.
В данной реакции карбоксилирование происходит с участием фермента рибулозодифосфаткарбоксилазы (РДФ-карбоксилаза).
Это самый распространенный в мире фермент.
Продукт реакции, содержащий 6 атомов углерода, в присутствии воды сразу распадается на две молекулы 3-фосфоглицириновой кислоты (3-ФГК):

С данной реакции и начинается цикл Кальвина.
ФГК и является, по современным взглядам, первичным продуктом ассимиляции углерода.
Для дальнейших превращений ФГК необходимы вещества световой фазы фотосинтеза: АТФ и НАДФН. Сначала 3-ФГК фосфорилируется при участии АТФ и образуется 1,3-дифосфоглицириновая кислота. Реакция катализируется ферментом фосфоглицераткиназой:

Затем происходит восстановление за счет НАДФН и образуется фосфоглицириновый альдегид ФГА:

Суммарный результат второй стадии: восстановление карбоксильной группы кислоты (–СООН) до альдегидной (–СНО).
Процесс превращения катализируется дегидрогеназой фосфоглициринового альдегида.
Дальнейшее превращение фосфоглициринового альдегида может происходить 4 путями.
ФГА частично с помощью триозофосфатизомеразы превращается в фосфодиоксиацетон (ФДА):

Это первый путь превращения ФГА.
Таким образом, в клетку поступают две найпростейшие формы сахаров: альдоза (ФГА) и кетоза (ФДА).
Это трехуглеродные сахара (триозосахара) с присоединенной к ним фосфатной группой содержат больше химической энергии, чем ФГК. Это первые углеводы, которые образуются при фотосинтезе.
С помощью альдолазы фосфодиоксиацетон (ФДА) соединяется с другой молекулой ФГА и образуется молекула фруктозо-1,6-дифосфата (ФДФ).

Это второй путь превращения ФГА.
Фруктозо-1,6-дифосфат дефосфорилируется и превращается во фруктозо-6-фосфат (Ф-6-Ф), что сопровождается накоплением в среде неорганического фосфата.
Фруктозо-6-фосфат в дальнейшем может выйти из цикла и использоваться для синтеза запасных форм углеводов: сахарозы, крахмала, других полисахаридов.
Однако ФГА (третий путь) может реагировать с эквимолярным количеством Ф-6-Ф, в результате образуются равные количества ксилулозо-5-фосфата и эритрозо-4-фосфата (транскетолаза).
Затем эритрозо-4-фосфат реагирует с равным количеством ФДА и образуется седагептулозо-1,7-дифосфат (альдолаза), которая фосфорилируется до седагептулозо-7-фосфата с участием седагептулозодифосфатазы.

Четвертый путь превращения ФГА связан с его реакцией с седагептулозо-7-фосфатом с образованием равных (эквимолярных) количеств рибозо-5-фосфата и ксилулозо-5-фосфата. Ксилулозо-5-фосфат эпимиризуется, а рибозо-5-фосфат изомерезуется до рибулозо-5-фосфата, последняя фосфорилируется за счет АТФ и образуется рибулозо-1,5-дифосфат – первичное соединение цикла Кальвина (акцептор СО2).
В этих реакциях тратится еще три молекулы АТФ.

Из приведенных реакций цикла Кальвина видно, что фотосинтез, являясь процессом запасания энергии, тем не менее, для своего существования требует затраты энергии.
В цикле Кальвина образование фруктозо-6-фосфата можно представить в виде следующего суммарного выражения:
6СО2 + 12НАДФН + 12Н+ + 18АТФ + 11Н2О →
фруктозо-6-фосфат + 12НАДФ+ + 18АДФ + 17Фн
18 молекул АТФ запасают около 140 ккал и 12 НАДФН –
Следовательно, поглощено около 755 ккал энергии. При этом в гексозах запасается около 670 ккал/моль. При таком балансе КПД составляет около 90 %.
10 % энергии растрачивается на поддержание цикла.
АТФ и НАДФН, которые образуются в световой стадии и используются на восстановление СО2, получили название ассимиляционной силы.
Цикл Кальвина подразделяют на три фазы:
– карбоксилирующую РДФ + СО2 → 2ФГК;
– восстановительную ФГК → ФГА;
– регинирующую ФГА → РДФ.
Каждая шестая молекула ФГА выходит из цикла и используется на синтез сахарозы или полисахаров, тогда как остальные 5 молекул через приведенные выше промежуточные реакции преобразуются в три молекулы рибулозо-1,5-дифосфата.
Так как первичный продукт цикла Кальвина – ФГК – содержит три атома углерода, то этот цикл получил название С3-цикла ассимиляции СО2. Упрощенную схему цикла можно представить в виде (рис.2.18):

Рис. 2.18. Упрощенная схема цикла Кальвина
Последовательность реакций на пути преобразования СО2 в сахар удалось выявить благодаря использованию радиоактивного углерода 14С и хромотографии на бумаге.
Описанный цикл восстановления СО2 до сахаров локализован в хлоропластах, так же как и биосинтез крахмала из образованных в них гексозофосфатов.
«Главный» же по количеству сахар, запасенный в растительной клетке – сахароза, – синтезируется уже вне хлоропласта: в слое цитоплазмы, прилегающем к наружной мембране этой органеллы. Сахароза синтезируется из Ф-6-Ф, образованной из ФГА и ФДА, которые в отличие от других сахаров цикла (пентоз и гексоз) легко транспортируются через мембраны хлоропластов.
Скорость цикла Кальвина зависит не только от количества образованных в световой стадии АТФ и НАДФН, но и от их соотношений.
Только соотношение 3АТФ и 2НАДФН обеспечивает активное восстановление углерода и запасание энергии.
Когда степень сопряжения работы ЭТЦ фотосинтеза с фотофосфорилированием мала, тогда интенсивность фотосинтеза, в первую очередь, может снизиться за счет уменьшения количества рибулозо-1,5-дифосфата, так как в этом случае будет лимитироваться фосфорилирование рибулоза-5-фосфата.
Кроме этого, в цикле при недостатке АТФ и НАДФН уменьшается возможность восстановления ФГК до триоз и поэтому одновременно со снижением интенсивности фотосинтеза среди ассимилятов (продуктов фотосинтеза) клетки увеличивается часть неуглеродных соединений.
Такое явление характерно, например, для растений, выращенных при слабом освещении.
ПУТЬ ПРЕВРАЩЕНИЕ УГЛЕРОДА – ТЕМНОВАЯ ФАЗА ФОТОСИНТЕЗА
План лекции:
1. С3-путь фотосинтеза (цикл Кальвина)
2. С4-путь фотосинтеза (цикл Хетча — Слэка)
3. Влияние условий на интенсивность процесса фотосинтеза
3.1.Коэффициент использования солнечной энергии.
3.3 Влияние содержания СО2 в воздухе
3.4 Влияние снабжения водой
3.5 Снабжение кислородом и интенсивность фотосинтеза
3.6 Влияние минерального питания
1. С3-путь фотосинтеза (цикл Кальвина)
Темновая фаза фотосинтеза— это совокупность биохимических реакций, в результате которых происходит усвоение растениями углекислого газа атмосферы (СО2) и образование углеводов. Ферменты, катализирующие темновые реакции растворены в строме. Если оболочки хлоропласта разрушить, то эти ферменты из стромы вымываются, в результате чего хлоропласты теряют способность усваивать СО2. Сущность темновых реакций процесса фотосинтеза была раскрыта благодаря исследованиям американского ученого Мелвина Кальвина. За эту работу в 1961 г. Кальвину с сотрудниками была присуждена Нобелевская премия. При этом успех экспериментов определялся широким применением новых методов исследования.
Первый метод, использованный Кальвином,— метод радиоактивного углерода. Радиоактивные изотопы по химическим свойствам практически не отличаются от стабильных. Принимая участие в реакциях, они как бы помечают те соединения, в которые входят. Скорость распада радиоактивных изотопов пропорциональна их количеству. Излучение, испускаемое ими в процессе разложения, может быть легко измерено. Все это создает возможность использования метода радиоактивных изотопов при изучении химизма фотосинтеза.
Второй метод — хроматография на бумаге. Если вещества, разогнанные на хроматограмме, содержат радиоактивные атомы, то их легко обнаружить с помощью радиоавтографии. Применяя указанные методы и короткие световые экспозиции можно определить, какие вещества и в какой последовательности образуются из 14 СО2. В качестве объекта исследований была взята зеленая водоросль хлорелла. После кратковременных экспозиций на свету в присутствии 14 СО2 растения фиксировались горячим спиртом. Спиртовой экстракт концентрировался, разделялся хроматографически и анализировался. Опыты показали, что через 5 с пребывания в атмосфере 14 СО2 на свету большая часть радиоактивного углерода сосредоточивается в трехуглеродном соединении — 3-фосфоглицериновой кислоте (3-ФГК). Кальвин выдвинул предположение, что в хлоропластах имеется какое-то вещество-акцептор, которое, взаимодействуя с СО2, образует фосфоглицериновую кислоту (акцептор + СО2 → ФГК). Для того чтобы установить природу акцептора, была проведена серия опытов с изменяющимися внешними условиями (смена света и темноты в присутствии и в отсутствие 14 СО2). Оказалось, что после выключения света содержание ФГК некоторое время продолжает расти. Одновременно наблюдалось быстрое исчезновение пятиуглеродного соединения — рибулезо-1,5-бифосфата (РБФ). Через 30 с РБФ не обнаруживался, а количество ФГК не изменялось. Вместе с тем на свету РБФ не исчезал, и его содержание оставалось постоянным. Иная картина наблюдалась в отсутствие СО,. В этом случае ни в темноте, ни на свету содержание РБФ не изменялось. Из полученных данных следовало, что в присутствии углекислого газа РБФ в темноте используется для образования ФГК. Дальнейшие превращения ФГК требуют света. В силу этого Кальвин выдвинул следующую предварительную схему процесса фотосинтеза:
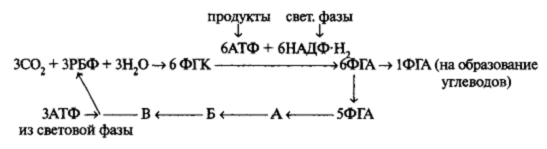 Согласно этой схеме РБФ является акцептором, который присоединяет СО2, в результате чего образуется ФГК. Однако в отсутствие света РБФ быстро оказывается использованным и исчезает. При этом накапливается известное количество ФГК. Именно это и наблюдалось в эксперименте. На свету при участии продуктов световой фазы происходит восстановление ФГК до фосфоглицеринового альдегида (ФГА). Судьба образовавшихся молекул ФГА различна. Частично путем ряда превращений ФГА используется на регенерацию акцептора (РБФ). Для процесса регенерации РБФ также необходима АТФ, образовавшаяся в световой фазе. В силу этого количество РБФ на свету поддерживается на постоянном уровне. В каждом цикле принимают участие три молекулы акцептора (РБФ) и образуется 6 молекул триозы (ФГА). Пять молекул ФГА идет на регенерацию акцептора через ряд промежуточных продуктов (на схеме А, Б, В). Каждая шестая молекула ФГА выходит из цикла и используется для построения углеводов и других метаболитов. Рассмотрим этот цикл более подробно.
Согласно этой схеме РБФ является акцептором, который присоединяет СО2, в результате чего образуется ФГК. Однако в отсутствие света РБФ быстро оказывается использованным и исчезает. При этом накапливается известное количество ФГК. Именно это и наблюдалось в эксперименте. На свету при участии продуктов световой фазы происходит восстановление ФГК до фосфоглицеринового альдегида (ФГА). Судьба образовавшихся молекул ФГА различна. Частично путем ряда превращений ФГА используется на регенерацию акцептора (РБФ). Для процесса регенерации РБФ также необходима АТФ, образовавшаяся в световой фазе. В силу этого количество РБФ на свету поддерживается на постоянном уровне. В каждом цикле принимают участие три молекулы акцептора (РБФ) и образуется 6 молекул триозы (ФГА). Пять молекул ФГА идет на регенерацию акцептора через ряд промежуточных продуктов (на схеме А, Б, В). Каждая шестая молекула ФГА выходит из цикла и используется для построения углеводов и других метаболитов. Рассмотрим этот цикл более подробно.
Цикл Кальвина можно разделить на фазы.
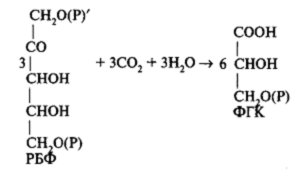
Первая фаза — карбоксилирование.Зга реакция катализируется специфическим для процесса фотосинтеза ферментом рибулозобисфосфат-карбоксилазой/ оксигеназой (сокращенно РБФ-карбоксилаза/оксигеназа), в научной литературе последних лет чаще встречается под названием Rubisco, от ribulosobiphosphat-ecarboxylase/oxygenase. Впервые Rubisco был выделен и очищен в 1955 году. Особенностью фермента является то, что катализируемаяимреакция является самой медленной стадией в цикле фиксации углекислоты. В листьях Rubisco содержится в больших количествах и является основной фракцией белка хлоропластов. Более того, высказывается мнение, что это самый распространенный белок-фермент на земном шаре. В активное состояние фермент переходит при освещении хлоропластов. Уже отмечалось, что формирование этого фермента происходит под контролем двух геномов: большие субъединицы (54 кДа) кодируются в ядре, синтезируются в цитоплазме; малые — кодируются и синтезируются в хлоропласте.
При взаимодействии РБФ с СО2 образуется сначала промежуточное нестойкое шестиуглеродное соединение, которое затем распадается на две молекулы ФГК.
Образовавшаяся ФГК — это органическая кислота, и ее энергетический уровень ниже уровня Сахаров. Поэтому это соединение не может непосредственно превращаться в углеводы. Необходимо превращение его в трехуглеродный сахар — фосфоглицериновый альдегид (ФГА).
Вторая фаза — восстановление.Дальнейшие превращения ФГК требуют участия продуктов световой фазы фотосинтеза: АТФ и НАДФН + Н + . Реакция идет в два этапа. Прежде всего, происходит реакция фосфорилирования 3-ФГК. Донором фосфатной группы является АТФ. АТФ требуется здесь в качестве дополнительного источника энергии. Образуется 1,3-дифосфоглицериновая кислота. Реакция катализируется ферментом фосфоглицерокиназой:
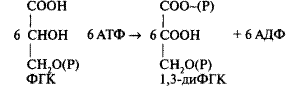
Образовавшееся в этой реакции соединение — дифосфоглицериновая кислота — обладает более высокой реакционной способностью, содержит макроэргическую связь, полученную от АТФ. Затем карбоксильная группировка этого соединения восстанавливается до альдегидной с помощью триозофосфатдегидрогена-зы, коферментом которойслужит НАДФ:
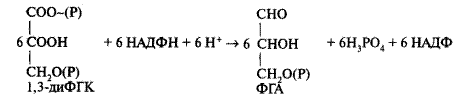
Образовавшийся ФГА является по уровню восстановленности углеводом. Это соединение вступает в две последние фазы. Пять молекул ФГА используется на регенерацию акцептора РБФ для того, чтобы фиксация СО2 могла снова осуществляться. Оставшаяся шестая молекула вступает в фазу «синтеза продуктов», где превращается в более сложные соединения (углеводы, аминокислоты и др.).
Третья фаза — регенерация.В процессе регенерации акцептора используется пять молекул ФГА, в результате чего образуются три молекулы рибулезо-5-фос-фата. Этот процесс идет через образование 4-, 5-, 6-, 7-углеродных соединений. Прежде всего, перваямолекула ФГА изомеризуется до фосфодиоксиацетона. Процесс катализируется ферментом триозофосфатизомеразой:

Фосфодиоксиацетон (ФДА) взаимодействует со второймолекулой ФГА с образованием фруктозодифосфата (ФДФ):
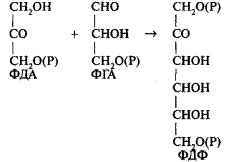
От ФДФ отщепляется фосфат, и ФДФ превращается во фруктозо-6-фосфат (ф-6-Ф). Далее от Ф-6-Ф (С6) отщепляется 2-углеродный фрагмент (—СО— —СН2ОН), который переносится на следующую (третью)триозу. Это транске-толазная реакция идет при участии фермента транскетолазы. В результате образуется первая пентоза (С5)-рибулезофосфат. От Ф-6-Фостается 4-углеродный сахар эритрозофосфат (С4), который конденсируется с четвертойтриозой с образованием седогептулезодифосфата (С7). После отщепления фосфата седогепту-лезодифосфат превращается в седогептулезофосфат. Далее снова происходит транскетолазная реакция, в результате которой от седогептулезофосфата отщепляется 2-углеродный фрагмент, который переносится на пятуютриозу. Образуются еще две молекулы рибулезофосфата. Таким образом, в результате рассмотренных реакций получаются 3 молекулы рибулезофосфата. Для образования из них акцептора (РБФ) необходимо их фосфорилирование. Для этого используются три молекулы АТФ, образовавшиеся в результате световых реакций. Все реакции, входящие в цикл Кальвина, представлены на схеме (рисунок 1).
Общее суммарное уравнение цикла следующее:
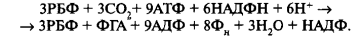
Образовавшийся триозофосфат (ФГА) вступает в четвертую стадию темно-вых реакций — стадию образования продуктов фотосинтеза.
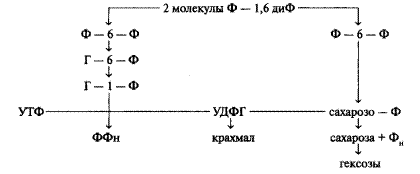
При прохождении двух циклов из 12 молекул образовавшегося ФГА две молекулы выходят из них, образуя одну молекулу фруктозодифосфата (ФДФ). Из двух молекул фруктозодифосфата (Ф-1, 6-диФ) образуются фруктозо-6-фосфат (ф-6-Ф) и глюкозо-1-фосфат (Г-1-Ф). Глюкозо-1-фосфат, взаимодействуя с ури-динтрифосфатом (УТФ), дает уридиндифосфоглюкозу (УДФГ).
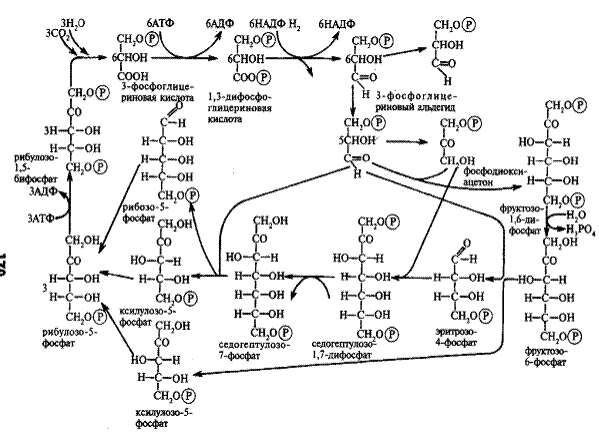
 Рисунок 1 — Путь углерода в фотосинтезе (цикл Кальвина)
Рисунок 1 — Путь углерода в фотосинтезе (цикл Кальвина)
В свою очередь УДФГ, реагируя с Ф-6-Ф, дает сахарозофосфат. Из сахарозофосфата путем дефосфорилирования образуется сахароза. Для образования одной молекулы сахарозы необходимо, чтобы прошли четыре цикла Кальвина:
По-видимому, именно сахароза является первым свободным сахаром, образующимся в процессе фотосинтеза. Из сахарозы образуются нефосфорилированные моносахара (глюкоза и фруктоза). Крахмал образуется из аденозиндифосфоглюкозы (ДЦФГ) или уридиндифосфоглюкозы (УДФГ), процесс катализируется ферментом амилосинтетазой.
Среди первых продуктов фотосинтеза обнаружены аминокислоты. По-видимому, ФГК, образовавшаяся на первом этапе цикла Кальвина, может превращаться в пировиноградную кислоту. Этот процесс идет особенно интенсивно при недостатке НДДФН, из-за чего задерживается преобразование ФГК в ФГА (обычный путь в цикле Кальвина). Пировиноградная кислота в присутствии NH, дает аминокислоту аланин. Показано, что скорость включения 14 СО2 в аланин в клетках хлореллы при некоторых условиях может даже превышать скорость ее включения в сахарозу. Из пировиноградной кислоты может образоваться еще ряд органических кислот (в цикле Кребса). Образовавшиеся органические кислоты в процессе аминирования или переаминирования дают аминокислоты. Сам по себе синтез аминокислот еще не означает образование белков. Однако было показано, что между этими двумя процессами имеется прямая связь. Так, под влиянием освещения синими лучами (458—480 нм) усиливается фотосинтетическое образование, как аминокислот, так и белков. В присутствии ингибиторов синтеза белка действие синего света не проявляется.
Наконец, из промежуточных продуктов цикла Кальвина могут образовываться жиры, липиды и другие продукты. Состав продуктов, образующихся при фотосинтезе, может быть определен исходя из величин фотосинтетического коэффициента. Под фотосинтетическим коэффициентомпонимается отношение выделенного в процессе фотосинтеза кислорода к поглощенному СО2. Если в процессе фотосинтеза образуются углеводы, то, согласно приведенному суммарному уравнению, фотосинтетический коэффициент должен быть равен единице: 6О2/6СО2 = 1. При образовании соединений, более восстановленных (содержащих меньше кислорода) по сравнению с углеводами, фотосинтетический коэффициент должен быть больше единицы. В случае образования белков фотосинтетический коэффициент равен 1,25, в случае жира — 1,44. Средняя величина фотосинтетического коэффициента для 27 видов растений оказалась равной 1,04. Расчеты показали, что такая величина фотосинтетического коэффициента указывает на образование наряду с углеводами некоторого количества белка (примерно 12%). Установлено, что величина фотосинтетического коэффициента меняется в зависимости от условий. Преобладание синих лучей над красными приводит к увеличению доли образующихся белков, тогда как красный свет благоприятствует образованию углеводов (Н.П. Воскресенская). Усиление снабжения растений азотом, естественно, также приводит к повышению фотосинтетического коэффициента и увеличению первичного синтеза белка. Имеются данные, что на качество продуктов фотосинтеза оказывает влияние интенсивность освещения. При высокой освещенности образуется больше углеводов, а при пониженной — аминокислот. Таким образом, изменяя условия среды, можно регулировать соотношение продуктов фотосинтеза. Указанные закономерности имеют большое не только теоретическое, но и практическое значение, так как позволяют направленно регулировать химический состав сельскохозяйственных растений и создавать условия для преимущественного синтеза углеводов, белков или жиров (А.А. Ничипорович). Выяснение механизма регуляции образования тех или иных продуктов фотосинтеза дает возможность улучшить состав сельскохозяйственных культур. Примером в этом отношении могут служить опыты с одноклеточной водорослью хлореллой, у которой удалось повысить содержание аминокислот и жиров за счет снижения содержания углеводов путем добавления в питательную среду NH4C1 (10
3 М). Все же основным продуктом фотосинтеза являются сахара. В связи с этим можно следующим образом расшифровать суммарное уравнение фотосинтеза.

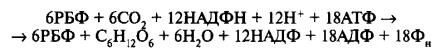
На основании приведенных реакций можно рассчитать энергетический баланс цикла Кальвина. Для восстановления шести молекул СО2 до уровня углеводов (глюкозы) требуется 18 молекул АТФ и 12 НАДФН. Соответственно для восстановления до уровня углеводов одной молекулы СО2 необходимы три молекулы АТФ и две НАДФН. Как мы видели, для образования двух молекул НАДФН и двух молекул АТФ необходимо 8 квантов света. Недостающее количество АТФ образуется в процессе циклического фотофосфорилирования. Следовательно, для восстановления одной молекулы СО2 до уровня углеводов должно быть затрачено 8—9 квантов. Энергия квантов красного света равна 168 кДж/моль. Таким образом, при использовании квантов красного света на восстановление одной молекулы СО, до уровня углеводов затрачивается примерно 1340—1508 кДж. Из этой энергии в у6 моль гексозы откладывается 478 кДж. КПД фотосинтеза в этом случае должен составить около 30—35%. Однако в естественных условиях коэффициент использования света значительно меньше.
В отличие от ферментов, принимающих участие в цепи переноса электронов (световая фаза фотосинтеза), ферменты цикла Кальвина локализованы в стро-ме хлоропластов. Согласованному осуществлению всех реакций способствует то, что эти ферменты часто ассоциированы на поверхности мембран и составляют определенные ансамбли.
Путь ассимиляции углерода при фотосинтезе, установленный Кальвином и предложенный в виде цикла Кальвина, является общим для всех автотрофных организмов. Однако существуют разные пути передачи СО2 в цикл Кальвина. Так, австралийские ученые М.Д. Хетч и К.Р. Слэк (1966) и советский ученый Ю.С. Карпилов (1960) показали, что у некоторых растений, преимущественно тропических и субтропических (в том числе кукуруза, сахарный тростник, амарант, просо, сорго), фотосинтез идет несколько по-иному.
В этом случае первым продуктом карбоксилирования является соединение, содержащее 4 атома углерода. Поэтому этот путь получил название С4-пути, в отличие от цикла Кальвина, в котором образуется ФГК, содержащая 3 атома углерода (С3-путь).
2. С4-путь фотосинтеза (цикл Хетча — Слэка)
Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по С4-пути, имеются два типа клеток и хлоропластов: 1) мелкие гранальные пластиды в клетках мезофилла листа; 2) крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки. Клетки обкладки имеют утолщенные клеточные стенки, содержат большое количество хлоропластов и митохондрий, расположены вокруг сосудистых пучков в 1 или 2 слоя. Совокупность указанных особенностей анатомического строения получила название корончатой анатомии или корончатого синдрома (от слова kranz — корона).
Хлоропласта разных типов клеток характеризуются не только особенностями строения, но и разным типам фосфорилирования. В клетках мезофилла по преимуществу происходит нециклическое фосфорилирование и образуется НАДФН, необходимый для цикла Кальвина, идущего в клетках обкладки. В хло-ропластах клеток обкладки идет только циклическое фосфорилирование. Такое разделение типов фосфорилирования, возможно, связано с тем, что к хлоро-пластам клеток обкладки, расположенным в глубине листа, проникает по преимуществу более длинноволновый свет, который не поглощается фотосистемой, ответственной за разложение Н2О.
На первом этапе С4-пути углекислый газ, диффундирующий в лист через устьица, попадает в цитоплазму клеток мезофилла с мелкими хлоропластами, в которых и происходит реакция карбоксилирования фосфоенолпировиноградной кислоты (ФЕП):
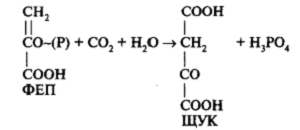
Реакция катализируется ферментом фосфоенолпируваткарбоксилазой (ФЕП-карбоксилазой) с образованием щавелевоуксусной кислоты (оксалоацетат). ЩУК преобразуется в яблочную кислоту (малат) или аспарагиновую (аспартат). Восстановление до маната происходит в присутствии НАДФН, а для образования аспартата необходимо наличие NH4 + . Затем яблочная (или аспарагиновая) кислота, по-видимому, по плазмодесмам передвигается в клетки обкладки. В клетках обкладки яблочная кислота декарбоксилируется ферментом малатдегидрогеназой до пировиноградной кислоты <пируват) и СО2. Реакция декарбоксилирования может варьировать у разных групп растений с использованием разных ферментов. СО2 поступает в хлоропласты клеток обкладки и включается в цикл Кальвина—присоединяется к РБФ (рисунок 2). Пируват возвращается в клетки мезофилла и превращается в первичный акцептор СО2 — ФЕП.
Таким образом, при С4-пути реакция карбоксилирования происходит дважды. Это позволяет растению создавать запасы углерода в клетках. Акцепторы Ор2 (ФЕП и РБФ) регенерируют, что и создает возможность непрерывного функционирования циклов. Фиксация СО2 с участием ФЕП и образованием малата или аспартата служит своеобразным насосом для поставки С02 в хлоропласты обкладки, функционирующих по С3-пути. Поскольку при таком механизме фотосинтеза принимают участие два типа клеток и два типа хлоропластов, этот путь называют еще кооперативным (Ю.С. Карпилов, 1970).
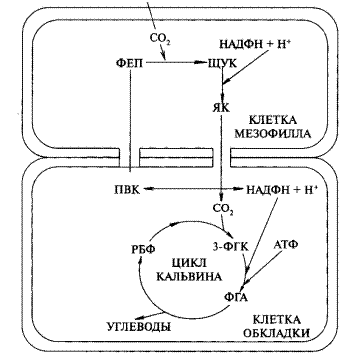
Рисунок 2 — Фотосинтетический цикл Хэтча-Саэка (С4 — путь):
ФЕП— фосфоенолпировиноградная кислота;
ЩУК— щавелевоуксусная кислота;
ЯК — яблочная кислота;
ПВК— пировиноградная кислота;
РБФ— рибулозо-1,5-бифосфат;
3-ФГК— фосфоглицериновая кислота;
ФГА— фосфоглицериновый альдегид
Высказывается мнение, что С4-путь возник в процессе эволюции как приспособление к изменившимся условиям среды. При возникновении фотосинтеза атмосфера была значительно богаче СО2 и беднее О2. Именно поэтому важнейший фермент цикла Кальвина Rubisco (РБФ-карбоксилаза/оксигеназа) может работать только при сравнительно высоких концентрациях СО2. Благодаря деятельности самих растений состав атмосферы изменился: содержание СО2 резко уменьшилось, а О2 возросло. В изменившихся условиях в осуществлении темновых реакций фотосинтеза появился ряд приспособительных черт. В частности значительно увеличилось содержание фермента Rubisco, который составляет почти половину белков стромы хлоропластов. Вместе с тем у некоторых растений выработался особый, дополнительный путь связывания СО2 с помощью ФЕП-карбоксилазы. Этот фермент обладает большим сродством к углекислоте и работает при концентрациях СО2 во много раз более низких по сравнению с Rubisco. Установлено, что и сопротивление мезофилла диффузии СО2 у С4-растений более чем в 3,5 раз меньше и составляет 0,3—0,8 см/с, в то время как у С3 — растений — 2,8 см/с.
Фиксация по С4-пути имеет еще ряд преимуществ. Растения С3-пути характеризуются высокой интенсивностью процесса, получившего название фотодыхания. Под фотодыханиемпонимают поглощение кислорода и выделение СО, на свету с использованием в качестве субстрата промежуточных продуктов цикла Кальвина. Как показали исследования, Rubisco (РБФ-карбоксилаза/оксигеназа) имеет двойственную функцию и может катализировать не только реакцию карбоксилирования цикла Кальвина: РБФ + СО2 →2ФГК. Rubisco способна реагировать с О2, осуществляя оксигеназную реакцию, при этом образуется фос-фогликолевая кислота:
РБФ + О2 → ФГК + фосфогликолевая кислота.
Фосфогликолевая кислота через ряд превращений распадается с выделением СО2. Таким образом, при фотодыхании часть промежуточных продуктов фотосинтеза теряется за счет выделения СО2. Реакции окисления и карбоксилирования конкурируют друг с другом, а осуществление Rubisco карбоксилазной или оксигеназной функции зависит от содержания О2 и СО2. Фотодыхание требует повышенной концентрации О2. Между тем, как уже упоминалось, в хлоропластах клеток обкладки концентрация О2 понижена, так как в них происходит только циклическое фосфорилирование, при котором вода не разлагается и О2 не выделяется. Вместе с тем в клетках обкладки повышена концентрация СО2. Такие условия ингибируют процесс фотодыхания в клетках обкладки и поэтому растения С4-типа характеризуются очень низкой потерей СО2 в результате фотодыхания.
Потери на фотодыхание у С3-растений особенно возрастают при повышении температуры и освещенности. В этой связи понятно, что растения С4 — это, главным образом, южные и даже тропические, которые получают дополнительные преимущества в смысле продуктивности фотосинтеза. Оптимум температуры для фотосинтеза у С3-растений 20—25 0 С, тогда как у растений С4 30—45 0 С.
Светонасыщение фотосинтеза С4-растений также происходит при более высоких значениях интенсивности света, чем у С3-растений. Так, у растений С3-пути интенсивность фотосинтеза перестает увеличиваться при 50% от полного солнечного освещения, в то время как у С4-форм этого не происходит. Такие особенности С4-растений объясняют высокую интенсивность фотосинтеза при повышенных температуре и освещенности. Характерным признаком растений С4-пути является, наконец, то, что образование продуктов цикла Кальвина происходит в хлоропластах, расположенных непосредственно около проводящих пучков. Это благоприятствует оттоку ассимилятов и, как следствие, повышает интенсивность фотосинтеза. Различия между С3— и С4-растениями можно продемонстрировать, поместив их рядом в одной камере (например, кукурузу и бобы) при высокой температуре и освещенности. Окажется, что СО2, выделяясь в процессе дыхания, постепенно переходит к кукурузе и соответственно изменяет темпы ее роста. Кукуруза как бы «съедает» растения бобов.
Показано, что С3-растения ассимилируют СО2 на полном солнечном свету со скоростью 1—50 мг/дм 2 ч, а С4-растения — со скоростью 40—80 мг/дм 2 ч. Кукуруза, сорго, просо, сахарный тростник являются одними из наиболее продуктивных культур. Так, интенсивность фотосинтеза у кукурузы 85 мг СО2/дм 2 -ч, сорго — 55 мг СО2/дм 2 -ч, тогда как у пшеницы всего 31 мг СО2/дм 2 ч. Высокая потенциальная продуктивность С4-растений наиболее полно реализуется при полном солнечном освещении и высокой температуре.
Важной физиологической особенностью С4-растений является их высокая засухо- и термоустойчивость. По мнению ряда исследователей возникновению С4-пути фотосинтеза способствовали засушливые условия окружающей среды. Уже отмечалось, что пространственное разделение процессов позволяет растениям с С4-путем фотосинтеза осуществлять фиксацию углекислоты даже приотносительно закрытых устьицах, поскольку хлоропласты клеток обкладки используют СО2, накопленный в виде доноров СО2 (малат или аспартат). Известно, что закрывание устьиц на наиболее жаркое время дня сокращает потери воды устьиц). Вместе с тем листья светолюбивых растений, а также верхние ярусы листьев, характеризуются большей толщиной, с сильно развитой палисадной паренхимой. В некоторых случаях палисадная паренхима располагается не только с верхней, но и с нижней стороны листа. Листья теневыносливых растений, как правило, имеют более крупные хлоропласты, с большим содержанием пигментов и несколько иным их соотношением. Уже отмечалось, что теневыносливые растения имеют больший размер светособирающего комплекса. Хлоропласты этих растений содержат относительно больше хлорофилла и ксантофилла по сравнению со светолюбивыми. Эти особенности в содержании состава пигментов позволяют листьям теневыносливых растений поглощать и использовать малые количества света, а также участки солнечного спектра, уже прошедшие через листья светолюбивых растений.
Важной особенностью, определяющей возможность растений произрастать при большей или меньшей освещенности, является положение компенсационной точки. Под компенсационной точкой понимается та освещенность, при которой процессы фотосинтеза и дыхания уравновешивают друг друга. Иначе говоря, это та освещенность, при которой растение за единицу времени образует в процессе фотосинтеза столько органического вещества, сколько оно тратит в процессе дыхания. Естественно, что рост зеленого растения может идти только при освещенности выше компенсационной точки. Чем ниже интенсивность дыхания, тем ниже компенсационная точка и тем при меньшей освещенности растения растут. Теневыносливые растения характеризуются более низкой интенсивностью дыхания, а соответственно и компенсационной точкой, что позволяет расти при меньшей освещенности. Компенсационная точка заметно растет с повышением температуры, так как повышение температуры сильнее увеличивает дыхание по сравнению с фотосинтезом. Именно поэтому при пониженной освещенности (например, в оранжереях зимой) необходима умеренная положительная температура; повышение температуры в этих условиях может снизить темпы роста растений. У ряда светолюбивых растений, таких, как кукуруза, просо, сорго, интенсивность фотосинтеза непрерывно возрастает и световое насыщение <выход на плато) не достигается даже при самой высокой освещенности. Для растений менее светолюбивых увеличение интенсивности освещения свыше 50% от полного солнечного освещения оказывается уже излишним. Для растений теневыносливых и особенно тенелюбивых (мхи, водоросли) выход на плато фотосинтеза происходит уже при 0,5— 1 % от полного дневного света.
3. Наконец, обсуждая вопрос об использовании света растениями, необходимо также подчеркнуть, что конечный выход продуктов фотосинтеза зависит от скорости не столько световых, сколько темновых реакций. В настоящее время показано, что свет оказывает стимулирующее влияние на работу ряда ферментов (Rubisco, АТФ-синтаза и др.). Активация этих ферментов под действием света связана с работой специального белка— тиоредоксина, содержащего тиоловые группы и способного к окислительно-восстановительным превращениям:
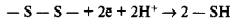
В хлоропластах тиоредоксин восстанавливается, принимая электроны от восстановленных молекул ферредоксина. Восстановленный тиоредоксин окисляется, отдавая, в свою очередь, электроны молекуле фермента. Таким образом, при переходе от темноты к свету, когда в хлоропластах начинает работать цепь переноса электронов, и образуются восстановленные молекулы ферредоксина, происходит активация Rubisco: ферредоксин — тиоредоксин — Rubisco .
Однако в основном с увеличением интенсивности освещения возрастает скорость световых реакций, и темновые реакции не успевают за ними. В этой связи снова необходимо обратить внимание на то, что темновые реакции пути С4 при высокой освещенности идут быстрее и меньше лимитируют использование продуктов световой фазы и, следовательно, общую интенсивность фотосинтеза. Возможно, это связано с тем, что активность фермента ФЕП-карбоксилазы более резко стимулируется светом по сравнению с Rubisco.
3. Влияние условий на интенсивность процесса фотосинтеза
3.1.Коэффициент использования солнечной энергии.
Показателем эффективности использования солнечной радиации растениями является коэффициент полезного действия (КПД). КПД— это отношение количества энергии, запасенной в продуктах фотосинтеза или образовавшейся в фитомассе урожая, к количеству поглощенной радиации:

где g — калорийность растений (ккал/г); у — биологический урожай общей сухой фитомассы (г/см 2 ); ∑Qф — сумма ФАРза вегетационный период (ккал/см 2 ). Под ФАР(фотосинтетически активной радиацией) понимается участок солнечного спектра, поглощаемый пигментами зеленого листа (380—740 нм). КПД выражается либо по отношению к падающей, либо по отношению к поглощенной растениями ФАР. Если рассматривать планету Земля в целом, то КПД падающей ФАР составляет около 0,2%. Следовательно, КПД фотосинтеза в естественных условиях ничтожно мал. Для разных растений и в разных условиях выращивания КПД поглощенной ФАР составляет следующие величины: кукуруза 2,5—5,7, ячмень 2,6—4,0, рис 2,5—4,4, озимая пшеница 1,1—6,3 (Х.Г. Туминг). Задача повышения КПД использования солнечной энергии является одной из важнейших в физиологии, а также в селекции сельскохозяйственных растений.
Влияние температуры на фотосинтез находится в зависимости от интенсивности освещения. При низкой освещенности фотосинтез от температуры не зависит (Q10 = 1). Следовательно, при низком уровне освещенности фотосинтез идет с одинаковой скоростью при 15° и 25°С. Этосвязано с тем, что при низкой освещенности интенсивность фотосинтеза лимитируется скоростью световых реакций. Напротив, при высокой освещенности скорость фотосинтеза определяется протеканием темновых реакций. В этом случае влияние температуры проявляется очень отчетливо и температурный коэффициент Q10 может быть около двух. Так, для подсолнечника повышение температуры в интервале от 9 до 19°С увеличивает интенсивность фотосинтеза в 2,5 раза. Температурные пределы, в которых возможно осуществление процессов фотосинтеза, различны для разных растений. Понижение температуры влияет на фотосинтез прямо, уменьшая активность ферментов, участвующих в темновых реакциях, и косвенно, благодаря повреждению органелл. Минимальная температура для фотосинтеза растений средней полосы около 0°С, для тропических растений 5—10°С. Имеются данные, что ранневесенние и высокогорные растения могут осуществлять фотосинтез и при температуре ниже 0°С. Лишайники могут ассимилировать СО2 при температуре —25°С. Оптимальная температура фотосинтеза для большинства растений средней полосы составляет примерно 20—25°С. При этом для растений, ведущих фотосинтез по С4-пути, оптимальная температура более высокая (35—45°С и выше), для цианобактерий оптимум температуры фотосинтеза значительно выше. При температуре выше оптимальной интенсивность фотосинтеза резко падает. Это связано с тем, что зависимость процесса фотосинтеза от температуры представляет собой равнодействующую противоположных процессов. Так, повышение температуры увеличивает скорость темновых реакций фотосинтеза. Одновременно при температуре 25—30°С происходит процесс инактивации хлоропластов. Повышение температуры может вызвать также закрытие устьичных щелей. Наконец, как уже говорилось, повышение температуры увеличивает интенсивность дыхания, и в этой связи видимый фотосинтез (разность между фотосинтезом и дыханием) уменьшается. Понижение температуры также снижает фотосинтез, поскольку тормозится активность ферментов, уменьшается скорость диффузионных процессов, а также отток ассимилятов.
3.3 Влияние содержания СО2 в воздухе
Источником углерода для процесса фотосинтеза является углекислый газ. Попытки заменить углекислый газ угарным (СО) не увенчались успехом. В основном в процессе фотосинтеза используется СО2 атмосферы. Правда, имеются данные, что частично СО2 может поступать в растения через корневую систему из почвы (АЛ. Курсанов). Однако этот источник имеет сравнительно малое значение.
Содержание СО2 в воздухе составляет всего 0,03%. Разные растения неодинаково используют одни и те же концентрации СО2. Растения, у которых фотосинтез идет по С4-пути (кукуруза, просо, сорго и др.), обладают более высокой способностью к связыванию СО2 благодаря высокой активности фермента ФЕП-карбоксилазы. Процесс фотосинтеза может осуществляться при содержании СО2 для C3-растений не менее 0,005, а для С4 — не менее 0,0005%.
Повышение содержания СО2 до 1,5% вызывает прямо пропорциональное возрастание интенсивности фотосинтеза у зерновых культур. Для других растений такое увеличение интенсивности фотосинтеза идет до 0,1%. При увеличении содержания СО2 до 15—20% процесс фотосинтеза выходит на плато, затем наступает депрессия фотосинтеза. Есть растения, более чувствительные к повышению концентрации СО2, у которых торможение фотосинтеза начинает проявляться уже при содержании СО2, равном 1%. Повышение концентрации СО2 оказывает ингибирующее влияние в силу разных причин. Увеличение содержания СО2 вызывает закрытие устьиц. Высокие концентрации СО2 сказываются особенно неблагоприятно при высокой освещенности. Это заставляет полагать, что СО2 в определенных концентрациях ингибирует отдельные ферментативные реакции цикла Кальвина. В естественных условиях содержание СО2 настолько мало, что может ограничивать возрастание процесса фотосинтеза. Особенно резко это проявляется при достаточно высокой интенсивности света, когда лимитирующими являются темновые реакции. Надо учесть, что в дневные часы содержание СО2, в воздухе вокруг растений понижается. Так, над полем кукурузы содержание СО2 днем снижается в 3 раза (до 0,01%). Это происходит несмотря на пополнение СО2 за счет перемещения слоев атмосферы и выделения СО2 в процессе дыхания (главным образом, почвенной микрофлорой).
В связи со сказанным увеличение содержания СО2 в воздухе является одним из важных способов повышения интенсивности фотосинтеза и, как следствие, накопления сухого вещества растением. Однако в полевых условиях регулирование содержания СО2 затруднено. Частично это может быть достигнуто с помощью усиления выделения СО2 почвой при внесении органических удобрений. Легче достигается повышение содержания СО2 в закрытом грунте. В этом случае подкормки СО2 дают хорошие результаты.
3.4 Влияние снабжения водой
Вода является непосредственным участником процесса фотосинтеза. Однако количество воды, необходимое для образования углеводов, ничтожно мало по сравнению с общим содержанием воды, необходимым для поддержания клетки в тургорном состоянии. Вместе с тем при полной насыщенности водой клеток листа фотосинтез снижается. Частично это может быть связано с тем, что при полном насыщении клеток мезофилла замыкающие устьичные клетки оказываются несколько сдавленными, устьичные щели не могут открыться (гидропассивные движения). Однако дело не только в этом. Небольшое обезвоживание листьев сказывается благоприятно на процессе фотосинтеза и вне зависимости от степени открытия устьиц. Так, погружение листьев в раствор маннита в концентрации 0,2—0,4 М вызывает водный дефицит, равный 15—20%, и некоторое возрастание интенсивности фотосинтеза. Таким образом, небольшой водный дефицит (5—15%) в клетках листьев оказывает благоприятное влияние на интенсивность фотосинтеза (В.А. Бриллиант). По-видимому, в процессе эволюции выработаны приспособления работы ферментов к небольшому водному дефициту. Действительно, небольшой водный дефицит активизирует ряд ферментов (Н.М. Сисакян).
Увеличение водного дефицита свыше 15—20% приводит к уже заметному снижению интенсивности фотосинтеза. Это связано в первую очередь с закрытием устьиц (гидроактивные движения), что резко уменьшает диффузию СО2 в лист. Кроме того, это вызывает сокращение транспирации, и, как следствие, возрастает температура листьев. Между тем повышение температуры выше 30°С вызывает снижение фотосинтеза. Наконец, обезвоживание оказывает влияние на конформацию, а следовательно, и активность ферментов, принимающих участие в темновой фазе фотосинтеза, повреждает структуру тилакоидов. При обезвоживании также замедляется процесс фотофосфорилирования (И.А. Тарчевский). Длительное обезвоживание растений может привести к тому, что интенсивность фотосинтеза не восстанавливается после улучшения снабжения водой.
3.5 Снабжение кислородом и интенсивность фотосинтеза
Несмотря на то, что кислород является одним из продуктов процесса фотосинтеза, в условиях полного анаэробиоза процесс фотосинтеза останавливается. Можно полагать, что влияние анаэробиоза косвенное, связано с торможением процесса дыхания и накоплением продуктов неполного окисления, в частности органических кислот, в связи с чем резко снижается значение рН. Это предположение подтверждается тем, что вредное влияние анаэробиоза сказывается более резко в кислой среде. Повышение концентрации кислорода (до 25%) также тормозит фотосинтез (эффект Варбурга).
Тормозящее влияние высоких концентраций кислорода на фотосинтез проявляется особенно резко при повышенной интенсивности света. Эти наблюдения заставили обратить внимание на особенности процесса дыхания в присутствии света. В осуществлении этого процесса принимают участие не только хлоропласты и митохондрии, но и особые органеллы — пероксисомы.
Как уже упоминалось, РБФ-карбоксилаза/оксигеназа может присоединить к РБФ не только СО2, но и О2. При естественной концентрации газов в атмосфере доля фотодыхания составляет 25—30%, поскольку способность связывать СО2 у фермента выше по сравнению с кислородом. При уменьшении концентрации СО2 и увеличении О2 доля фотодыхания возрастает. При отсутствии СО2 фотодыхание превышает фотосинтез. Химизм этого процесса отличен от обычного темнового дыхания. В результате присоединения кислорода РБФ распадается на одну молекулу ФГК и одну молекулу фосфогликолевой кислоты (фосфогликолат). Фосфогликолевая кислота путем дефосфорилирования превращается в гликолевую кислоту (гликолат). Образование гликолевой кислоты происходит в хлоропластах, однако она там не накапливается, а транспортируется в перок-сисомы (рисунок 3). В пероксисомах происходит превращение гликолевой кислоты

Рисунок 3 — Фотодыхание и метаболизм гликолевой кислоты
в глиоксилевую кислоту (глиоксилат). Образующаяся при этом перекись водорода расщепляется содержащейся в пероксисомах каталазой. Глиоксилевая кислота, в свою очередь, подвергается аминированию с образованием аминокислоты глицина. Глицин поступает в третий тип органелл — митохондрии, где 2 молекулы глицина образуют молекулу аминокислоты — серина и при этом выделяется СО2:
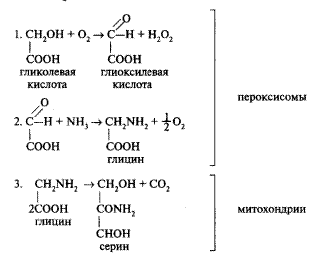
Не всегда гликолатный путь завершается в митохондриях. В некоторых случаях может замыкаться цикл. При этом серии поступает в пероксисому, где дезаминируется (отщепляется аминогруппа) с образованием гидроксипирови-ноградной кислоты (гидроксипируват). Гидроксипируват восстанавливается до глицерата, который в хлоропластах фосфорилируется с образованием 3-ФГК. Фосфоглицериновая кислота включается в цикл Кальвина.
Важно заметить, что при фотодыхании накопления энергии в АТФ не происходит. Выделение СО2 при фотодыхании С3-растений может достигать 50% от всего СО2, усвоенного в процессе фотосинтеза. Это особенно проявляется в условиях повышенного содержания О2, так как О2 и СО2 конкурируют за присоединение к РБФ. В связи с этим уменьшение интенсивности фотодыхания должно привести к повышению продуктивности растений. Так, мутантные формы табака, не обладающие способностью к образованию гликолевой кислоты, отличаются повышенным накоплением сухой массы. Имеются данные, что некоторое уменьшение содержания кислорода в атмосфере сказывается благоприятно на темпах накопления сухого вещества проростками. У кукурузы и других растений, осуществляющих фотосинтез по С4-пути, потери на фотодыхание минимальны. Не исключено, что такой тип обмена способствует большей продуктивности этих растений. Вместе с тем нельзя исключить, что процесс фотодыхания имеет определенное биологическое значение, в частности он способствует образованию аминокислот.
В настоящее время роль фотодыхания активно обсуждается. Существует мнение, что осуществление фотодыхания позволяет предотвратить деструкцию фотосинтетического аппарата, которая может происходить в отсутствие углекислого газа при невостребованности продуктов световой фазы. Реакции превращения гликолата позволяют обойти реакцию карбоксил ирования в цикле Кальвина, при этом цикл может функционировать без поглощения СО2. Кроме того, фотодыхание усиливается, когда образуется много углеводных продуктов фотосинтеза, а их экспорт из листа к потребляющим органам затруднен. В этих условиях образуются аминокислоты, которые расходуются на’синтетические процессы. Существует и такое мнение, что фотодыхание является формой приспособления растений к современному содержанию СО2 и О2 в атмосфере. Дело в том, что концентрация О2 превышает потребности растений для обеспечения темно-вого дыхания. Поэтому использование кислорода в процессе фотодыхания защищает растения от тормозящего влияния О2 на физиологические процессы клетки и целостность органелл.
3.6 Влияние минерального питания
Естественно, что исключение любого элемента минерального питания скажется на интенсивности фотосинтеза. Однако ряд элементов играет важную специфическую роль. Очень велико значение фосфорадля фотосинтеза. На всех этапах фотосинтеза принимают участие фосфорилированные соединения. Энергия света аккумулируется в фосфорных связях. При дефиците фосфора нарушаются фотохимические и темновые реакции фотосинтеза. Процессы фотофосфори-лирования требуют также обязательного присутствия магния.Имеются данные, что при недостатке калияинтенсивность фотосинтеза снижается уже через короткие промежутки времени. Калий может влиять на фотосинтез косвенно, через повышение оводненности цитоплазмы, ускорение оттока ассимилятов из листьев, увеличение степени открытия устьиц. Имеет место и прямое влияние калия, поскольку он активирует процессы фосфорилирования. При изучении фотосинтеза штамма хлореллы, который может расти как в темноте за счет готового органического вещества, так и на свету, было показано, что марганецнеобходим только в последнем случае. Отсутствие марганца резко угнетает реакцию Хилла и процесс нециклического фотофосфорилирования. Все это доказывает, что роль марганца определяется его участием в реакциях фотоокисления воды. Имеются данные, показывающие необходимость хлорадля фоторазложения воды. В процессе фотосинтеза участвуют многочисленные белки-ферменты. В этой связи понятно большое значение уровня азотного питания.Показано, что внесение азотных удобрений вызывает повышение фотосинтетической деятельности растений. Многие соединения, функционирующие как переносчики, содержат железо(цитохромы, ферредоксин) или медь(пла-стоцианин). Естественно, что при недостатке этих элементов интенсивность фотосинтеза понижается.
Фотосинтез
Типы питания
По типу питания живые организмы делятся на автотрофы, гетеротрофы и миксотрофы. Автотрофы (греч. αὐτός — сам + τροφ — пища) — организмы, которые самостоятельно способны синтезировать органические вещества из неорганических. Гетеротрофы (греч. ἕτερος — иной + τροφή — пища) — организмы, использующие для питания готовые органические вещества.
Наконец, миксотрофы (греч. μῖξις — смешение + τροφή — пища) — организмы, которые могут использовать как гетеротрофный, так и автотрофный способ питания. К примеру, эвглена зеленая на свету начинает фотосинтезировать, а в темноте питается гетеротрофно.

Фотосинтез
Фотосинтез (греч. φῶς — свет и σύνθεσις — синтез) — сложный химический процесс преобразования энергии квантов света в энергию химических связей. В результате фотосинтеза происходит синтез органических веществ из неорганических.

Этот процесс уникален и происходит только в растительных клетках, а также у некоторых бактерий. Фотосинтез осуществляется при участии хлорофилла (греч. χλωρός — зелёный и φύλλον — лист) — зеленого пигмента, окрашивающего органы растений в зеленый цвет. Существуют и другие вспомогательные пигменты, которые вместе с хлорофиллом выполняют светособирающую или светозащитную функции.
Ниже вы увидите сравнение строения хлорофилла и гемоглобина. Обратите внимание, что в центре молекулы хлорофилла находится ион Mg.
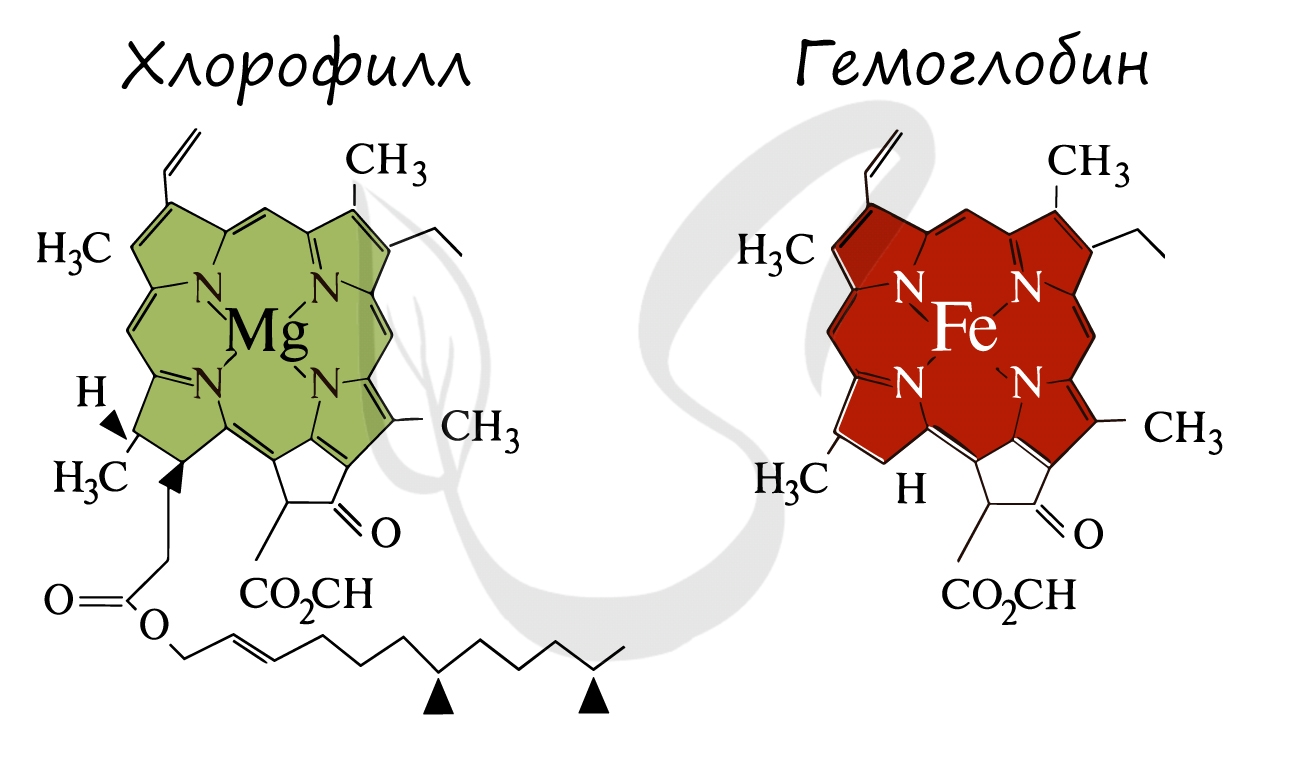
В высшей степени гениально значение процесса фотосинтеза подчеркнул русский ученый К.А. Тимирязев: «Все органические вещества, как бы они ни были разнообразны, где бы они ни встречались, в растении ли, в животном или человеке, прошли через лист, произошли от веществ, выработанных листом. Вне листа или, вернее, вне хлорофиллового зерна в природе не существует лаборатории, где бы выделялось органическое вещество. Во всех других органах и организмах оно превращается, преобразуется, только здесь оно образуется вновь из вещества неорганического»
Более подробно мы обсудим значение фотосинтеза в завершение этой статьи. Фотосинтез состоит из двух фаз: светозависимой (световой) и светонезависимой (темновой). Я рекомендую использовать названия светозависимая и светонезависимая, так как они способствуют более глубокому (и правильному!) пониманию фотосинтеза.
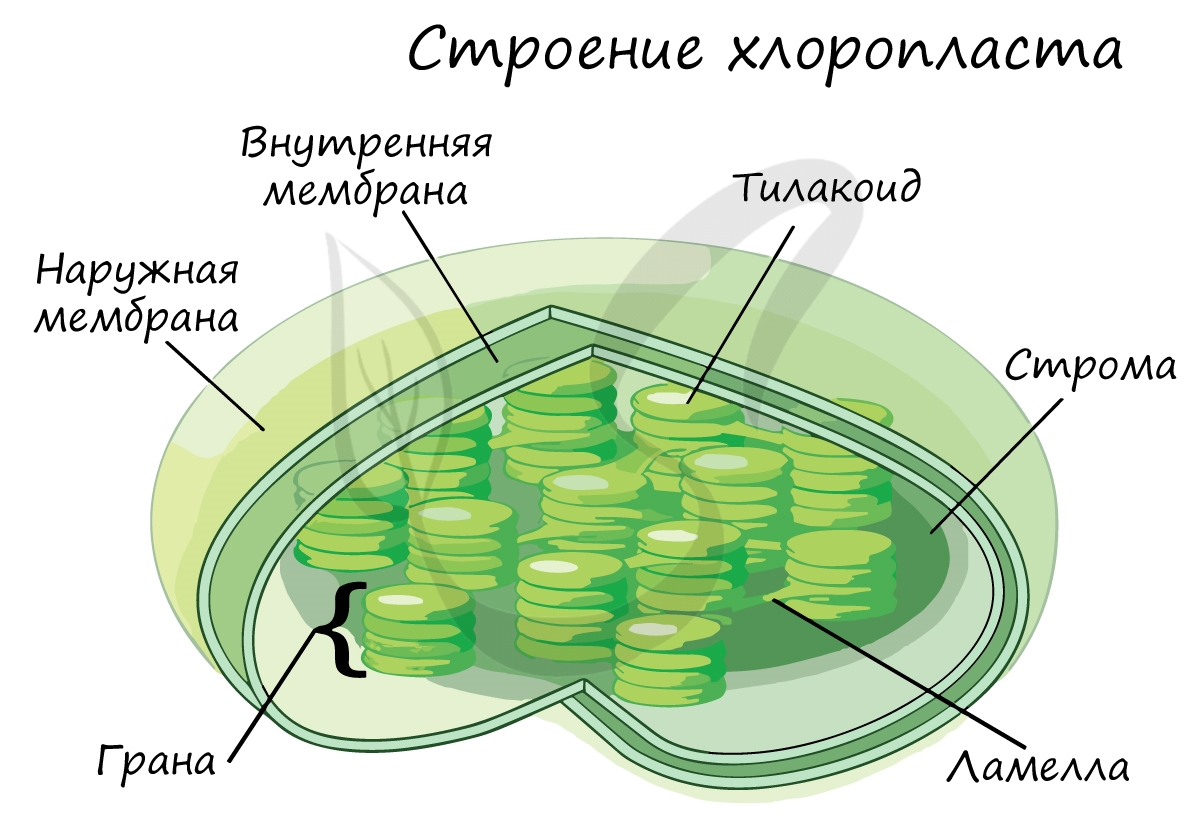
Светозависимая фаза (световая)
Эта фаза происходит только на свету на мембранах тилакоидов в хлоропластах. В ней принимают участие различные ферменты, белки-переносчики, молекулы АТФ-синтетазы и зеленый пигмент хлорофилл.
Хлорофилл выполняет две функции: поглощения и передачи энергии. При воздействии кванта света хлорофилл теряет электрон, переходя в возбужденное состояние. С помощью переносчиков электроны скапливаются с наружной поверхности мембраны тилакоидов, тем временем внутри тилакоида происходит фотолиз воды (разложение под действием света):
Гидроксид-ионы отдают лишний электрон, превращаясь в реакционно способные радикалы OH, которые собираются вместе и образуют молекулу воды и свободный кислород (это побочный продукт, который в дальнейшем удаляется в ходе газообмена).
Образовавшиеся при фотолизе воды протоны (H + ) скапливаются с внутренней стороны мембраны тилакоидов, а электроны — с внешней. В результате по обе стороны мембраны накапливаются противоположные заряды.
При достижении критической разницы, часть протонов проталкивается на внешнюю сторону мембраны через канал АТФ-синтетазы. В результате этого выделяется энергия, которая может быть использована для фосфорилирования молекул АДФ:
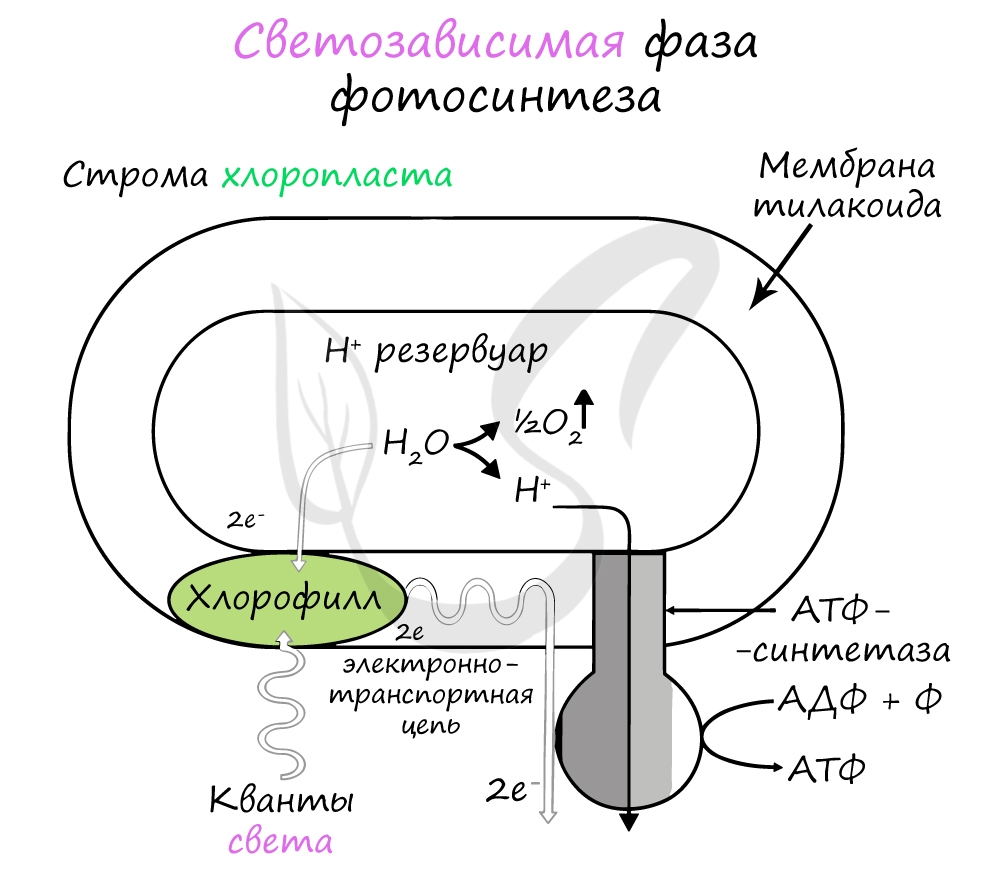
Протоны, попав на поверхность мембраны тилакоидов, соединяются с электронами и образуют атомарный водород, который используется для восстановления молекулы-переносчика НАДФ (никотинамиддинуклеотидфосфат). Благодаря этому окисленная форма — НАФД + превращается в восстановленную — НАДФ∗H2.
Предлагаю создать квинтэссенцию из полученных нами знаний. Итак, в результате светозависимой фазы фотосинтеза образуются:
- Свободный кислород O2 — в результате фотолиза воды
- АТФ — универсальный источник энергии
- НАДФ∗H2 — форма запасания атомов водорода
Кислород удаляется из клетки как побочный продукт фотосинтеза, он совершенно не нужен растению. АТФ и НАДФ∗H2 в дальнейшем оказываются более полезны: они транспортируются в строму хлоропласта и принимают участие в светонезависимой фазе фотосинтеза.
Светонезависимая (темновая) фаза
Светонезависимая фаза происходит в строме (матриксе) хлоропласта постоянно: и днем, и ночью — вне зависимости от освещения.
При участии АТФ и НАДФ∗H2 происходит восстановление CO2 до глюкозы C6H12O6. В светонезависимой фазе происходит цикл Кальвина, в ходе которого и образуется глюкоза. Для образования одной молекулы глюкозы требуется 6 молекул CO2, 12 НАДФ∗H2 и 18 АТФ.
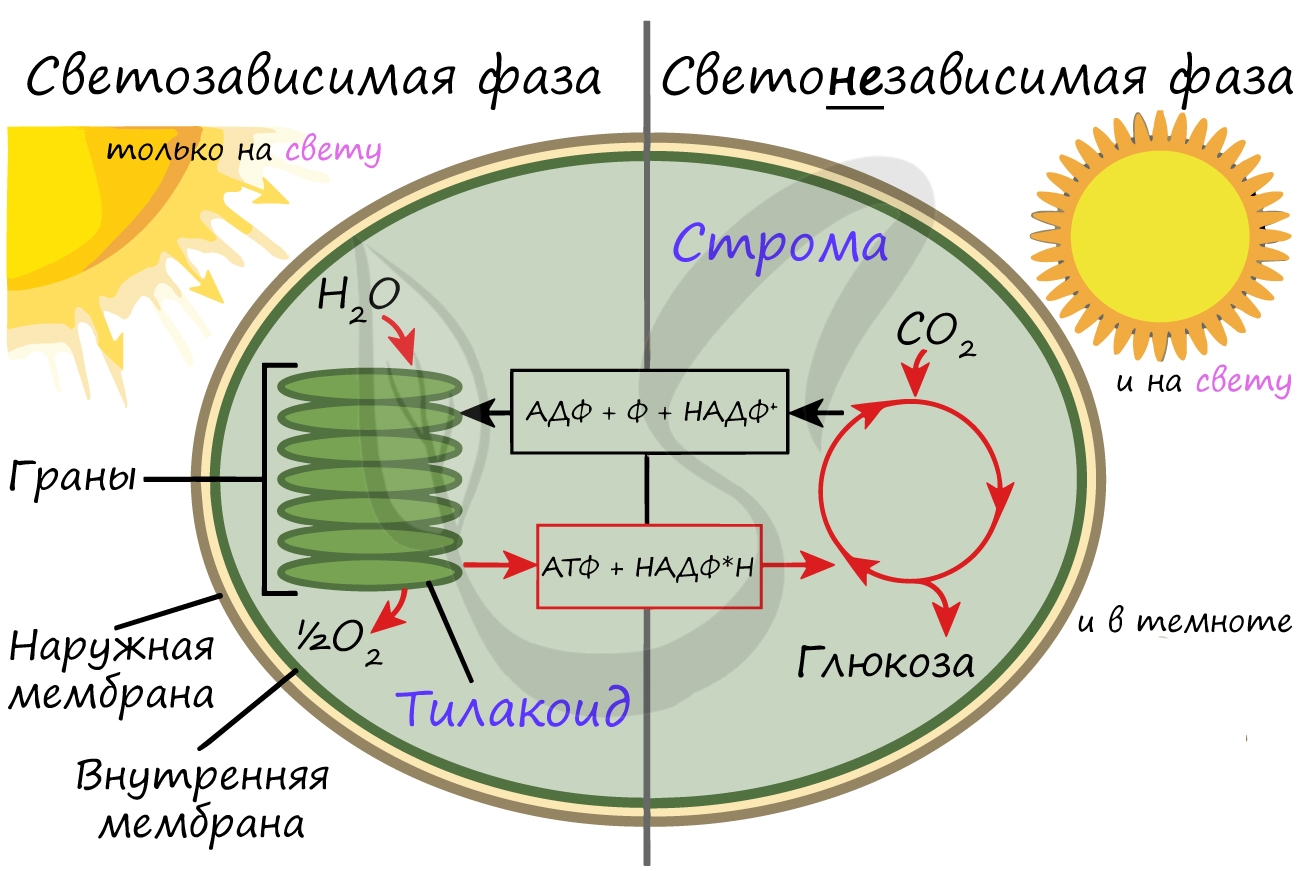
Таким образом, в результате темновой (светонезависимой) фазы фотосинтеза образуется глюкоза, которая в дальнейшем может быть преобразована в крахмал, служащий для запасания питательных веществ у растений.
Значение фотосинтеза
Значение фотосинтеза невозможно переоценить. Уверенно утверждаю: именно благодаря этому процессу жизнь на Земле приобрела такие чудесные и изумительные формы, какие мы видим вокруг себя: удивительные растения, прекрасные цветы и самые разнообразные животные.
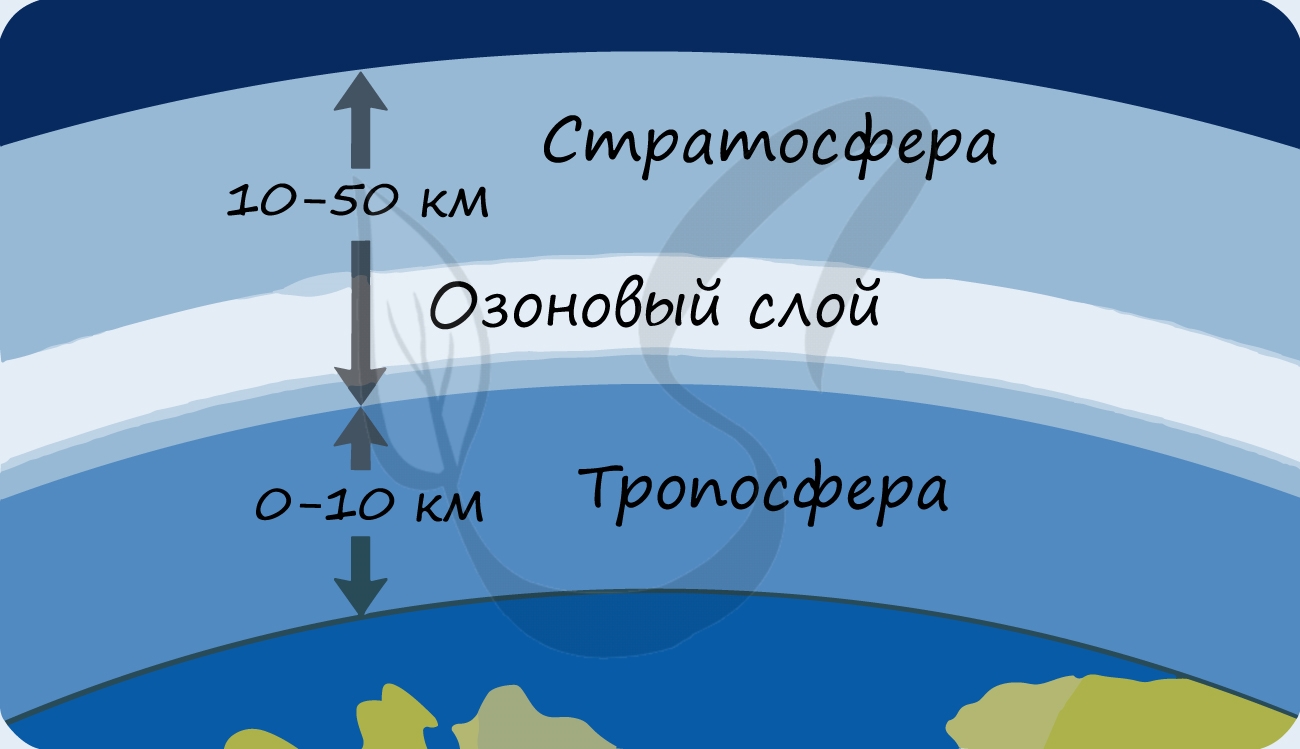
В разделе эволюции мы уже обсуждали, что изначально в составе атмосферы Земли не было кислорода: миллиарды лет назад его начали вырабатывать первые фотосинтезирующие бактерии — сине-зеленые водоросли (цианобактерии). Постепенно кислород накапливался, и со временем на Земле стало возможно аэробное (кислородное) дыхание. Возник озоновый слой, защищающий все живое на нашей планете от губительного ультрафиолета.
Говоря о роли фотосинтеза, выделим следующие функции, объединяющиеся в так называемую космическую роль растений. Итак, растения за счет фотосинтеза:
- Синтезируют органические вещества, являющиеся пищей для всего живого на планете
- Преобразуют энергию света в энергию химических связей, создают органическую массу
- Растения поддерживают определенный процент содержания O2 в атмосфере, очищают ее от избытка CO2
- Способствуют образованию защитного озонового экрана, поглощающего губительное для жизни ультрафиолетовое излучение

Хемосинтез (греч. chemeia – химия + synthesis — синтез)
Хемосинтез — автотрофный тип питания, который характерен для некоторых микроорганизмов, способных создавать органические вещества из неорганических. Это осуществляется за счет энергии, получаемой при окислении других неорганических соединений (железо- , азото-, серосодержащих веществ).
Хемосинтез был открыт русским микробиологом С.Н. Виноградским в 1888 году. Большинство хемосинтезирующих бактерий относится к аэробам, для жизни им необходим кислород.
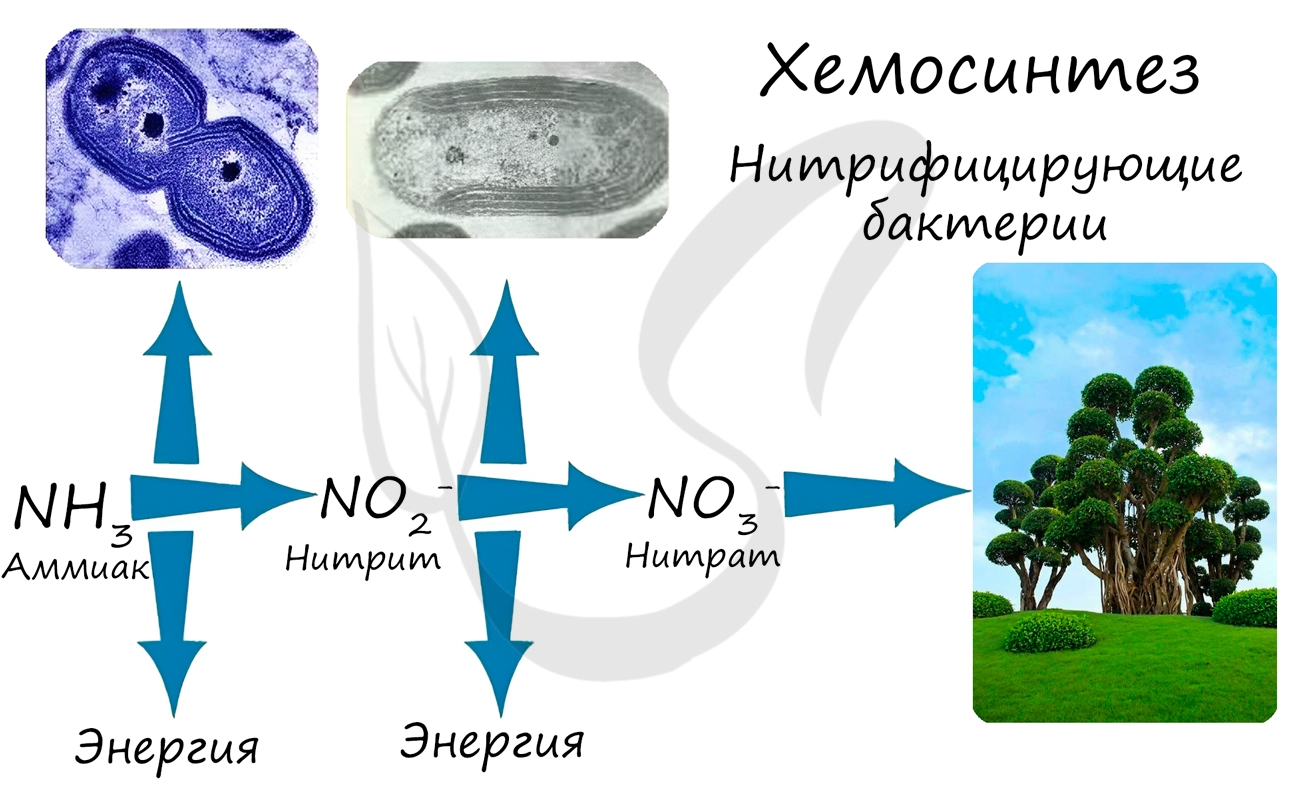
При окислении неорганических веществ выделяется энергия, которую организмы запасают в виде энергии химических связей. Так нитрифицирующие бактерии последовательно окисляют аммиак до нитрита, а затем — нитрата. Нитраты могут быть усвоены растениями и служат удобрением.
Помимо нитрифицирующих бактерий, встречаются:
- Серобактерии — окисляют H2S —> S 0 —> (S +4 O3) 2- —> (S +6 O4) 2-
- Железобактерии — окисляют Fe +2 —>Fe +3
- Водородные бактерии — окисляют H2 —> H +1 2O
- Карбоксидобактерии — окисляют CO до CO2
Значение хемосинтеза
Хемосинтезирующие бактерии являются неотъемлемым звеном круговорота в природе таких элементов как: азот, сера, железо.
Нитрифицирующие бактерии обеспечивают переработку (нейтрализацию) ядовитого вещества — аммиака. Они также обогащают почву нитратами, которые очень важны для нормального роста и развития растений.
Усвоение нитратов происходит за счет клубеньковых бактерий на корнях бобовых растений, однако важно помнить, что клубеньковые (азотфиксирующие) бактерии, в отличие от нитрифицирующих бактерий, питаются гетеротрофно.
© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.