Признак– это любое фенотипически проявляемое свойство организма. Большинство признаков являются генетически обусловленными вариациями, но их выражение может зависеть и от воздействия внешней среды (модификационные признаки). В зависимости от типа генетического контроля различают признаки качественные, количественные, пороговые и др.
Качественныепризнаки, как правило, контролируются одним геном, у которых наблюдается прерывистая изменчивость, описываемая по принципу “есть-нет”, например высокорослый — низкорослый, окрашенный – бесцветный, остистый – безостый.
Признаки количественные (полигенные, мерные)– это признаки, контролируемые суммарным действием большого числа генов, т.е. цифровое выражение результатов измерений, взвешиваний, подсчетов и т.д.
Полигенный контроль и большая вариабельность количественных признаков вследствие действия внешней среды обуславливают их непрерывную изменчивость по принципу “больше-меньше”, т.е. один и тот же признак, присущий разным особям или формам, имеет различную степень выражения.
Пороговые признаки – это определенные полигенные признаки проявляющиеся только при аддитивном (суммарном) действии соответствующих аллелей или при достижении их пороговой величины экспрессии.
К количественным признакам относят хозяйственно ценные (живая масса, удой, настриг шерсти, воспроизводительная способность, скорость роста и т. д.), а так же физиологические признаки. Они характеризуются типичным непрерывным изменением уровня у особей конкретной группы, определяемая как целыми, так и долями единиц их измерения.
К количественным признакам относят также и те, которые имеют прерывистое выражение, например, яйценоскость, плодовитость, ряд физиологических отличий.
В практике животноводства и ветеринарии, в научных исследованиях количественные признаки непрерывного и прерывистого типов изменчивости имеют важное значение, что предопределяет изучение генетических особенностей и закономерностей их изменения.
Большинство хозяйственно-полезных признаков домашних животных относится к группе количественных и имеет постоянно промежуточный тип наследования. Любой количественный признак развивается под влиянием наследственности и различных факторов среды.
Доля участия наследственой изменчивости в фенотипической изменчивости признака называется наследуемостью.
Наследуемостъ характеризуется с помощью коэффициента наследуемости (h 2 ). Абсолютное значение коэффициента наследуемости находится в пределах от 0 до 1.
Степень наследуемости признака можно охарактеризовать также с помощью коэффициента повторяемости (гw).
Повторяемостью называют способность организма удерживать свои показатели при постоянных условиях и сохранять свой ранг по сравнению с другими животными при изменении условий среды.
Показатели наследуемости используются для определения эффективности отбора в популяции.
Одним из основных показателей, выражающих изменения количественных признаков под влиянием селекции, служит величина селекционного эффекта (R), который показывает эффективность отбора (ответ на отбор).
Он выражается путем сравнения средних величин признака в двух смежных поколениях до и после отбора:
R=X потомства отобранных животных — Х популяция
то есть разница в уровне средней величины признака у потомства, полученного от отобранных родителей, и средним уровнем признака в популяции до отбора родителей.
Если сравнить средний уровень признака у отобранной лучшей группы родителей (X отоб. .род.) со средним уровнем признака в популяции (X попул.), то разность между ними составит так называемый селекционный дифференциал:
Соотношение между величинами R и Sd определено коэффициентом наследуемости h 2 , а именно: R=h 2 •Sd.
Следовательно, чем больше коэффициент наследуемости признака и селекционный дифференциал, тем выше эффект селекцииR,выявляемый у потомства отобранных родителей.
Соотношение селекционного дифференциала (Sd) и фенотипической изменчивости признака (Gp) выражает интен-сивность селекции i, а именно: i=Sd:σp. Следовательно, Sd=i*σp.
Используя полученное выражение, можно определить теоретический, то есть ожидаемый эффект селекции: R=i / σp*h’ 2 . Из этого выражения следует, что чем больше интенсивность отбора и коэффициент наследуемости, тем выше эффект селекции.
Интенсивность отбора определяется численностью исходного материала, то есть чем меньше доля отобранных для размножения животных, тем выше интенсивность селекции и выше эфффективность отбора (R).
Повышение коэффициента наследуемости достигается оптимизацией кормления и содержания, что уменьшает вариансу среды (σ 2 P) и увеличивает долю генетической вариансы (σp 2 ) в фенотипической изменчивости признака. Это, в свою очередь, повышает h 2 и эффект селекции.
Коэффициент повторяемости (rw) используют для прогнозирования количественного признака, оценки животных в ран-нем возрасте. Чем больше величина rw , то есть чем он ближе к единице, тем выше значение этого коэффициента для суждения о доли влияния наследственности на изменчивость признака и его наследуемость.
Генетический коэффициент корреляции (rG) позволяет прогнозировать при отборе по одному признаку возможный уровень другого. Он показывает, как оба селекционируемых признака изменяются при проведении отбора по одному из них или при отборе по двум. Зная величину rG, можно по одному признаку планировать или прогнозировать другой признак. Чем больше величина rG между двумя признаками, тем эффективнее косвенная селекция, если отбор ведут по одному признаку, а эффект проявляется и по другому, коррелирующему с основным признаком отбора.
Занятие 1. Генетика количественных признаков
Вид занятия: лабораторное
Цель занятия: Научиться вычислять коэффициенты наследуемости и повторяемости, определить эффект селекции и целевой стандарт для определенного стада.
Материальное обеспечение: практикум, индивидуальные задания, таблицы.
Содержание и методика проведения занятия.
Задание 1. Вычислите коэффициент корреляции (г) между признаками и определите коэффициент наследуемости (h 2 ).
Лимиты по признаку х = К для ряда х =
Лимиты по признаку у = К для ряда у =
 х у х у | fy | ay | fyay | fya 2 y |
| fx | ||||
| ах | ||||
| fхaх | ||||
| fхa 2 х |




Задание 2.Определите коэффициент повторяемости = rw (по удою, содержанию жира в молоке и т.д.)
| Лактация по счету | Порядковые номера коров |
| 3-я |




 = ;
= ;
= ;
 =
=
 =
=


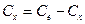
Далее расчеты ведутся по таблице дисперсионного анализа
| Источники варьирования | Сумма квадратов | Число степеней свободы | Средний квадрат | F факт. | F табл. |
| Общая изменчивость | C | δ у 2 | 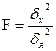 | F0,95= | |
| Между группами | C | δ х 2 | F0,95= | ||
| Внутри групп | C | δ z 2 |

Vx = число групп – 1 =ё

Vz = n – число групп



Определяем коэффициент повторяемости
Задание 3. Использование в селекции коэффициента наследуемости. Вычисление эффекта селекции и целевого стандарта.
Для определения эффекта селекции необходимо иметь следующие показатели (по удою или % жира и т.д.).
 — стада (ст.)
— стада (ст.)
— селекционного ядра (с.я.)
— матерей отцов (м.о.)
h 2 – по матерям
План ежегодной выбраковки (%)
Эффект селекции (Э.С.) определяется по формуле:

Sd – селекционный дифференциал
Sd – матерей = с.я.— ст.
Sd – отцов = м.о.— с.я.
Целевой стандарт определяется по формуле ст.+Э.С.
Э.С. за 1 год зависит от интервала между поколениями, который связан с процентом браковки. При ежегодной браковке 20% животных.

ст.= hм 2 =
с.я.= hо 2 =
м.о.= браковка %
Задание 4. Заполните таблицу значениями коэффициентов наследуемости по основным признакам и сделайте выводы об эффективности отбора по этим признакам в зависимости от величины h 2 .
- Глава 12. Генетика количественных признаков
- Лекция № 17. Основные понятия генетики. Законы Менделя
- Основные понятия генетики
- Методы генетики
- Генетическая символика
- Закон единообразия гибридов первого поколения, или первый закон Менделя
- Закон расщепления, или второй закон Менделя
- Закон чистоты гамет
- Цитологические основы первого и второго законов Менделя
- Закон независимого комбинирования (наследования) признаков, или третий закон Менделя
- Цитологические основы третьего закона Менделя
Глава 12. Генетика количественных признаков

ГЛАВА 12. ГЕНЕТИКА КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ
Большинство признаков и свойств организмов характеризуются количественным типом индивидуальной изменчивости, для которой типично непрерывное изменение величины признака у особей какой-либо группы. Величина количественного признака варьирует от минимального уровня у части особей к среднему — у других и далее к максимальному уровню у остальных. Даже в пределах достаточно однородной по полу, возрасту, породе группы животных у близкородственных особей наблюдается индивидуальная изменчивость признака, величину которого можно измерить. К количественным признакам относят хозяйственно ценные (живая масса, величина, удой, настриг шерсти) и физиологические признаки. Они характеризуются типичным непрерывным изменением уровня у особей конкретной группы. К количественным признакам относят также и те, которые имеют прерывистое выражение, например яйценоскость, плодовитость, а также ряд физиологических отличий.
Количественные признаки непрерывного и прерывистого типов изменчивости имеют важное значение в практике животноводства и ветеринарии, в научных исследованиях, поэтому необходимо изучать генетические особенности и закономерности их изменчивости.
Генетические основы наследования количественных признаков. Наследование количественных признаков обусловлено одинаковым или сходным действием многих доминантных неаллельных генов на признак (полимерия) либо многими однозначными генами (полигения). На наличие двух или трех пар однозначно действующих полимерных генов, определяющих степень выраженности признака, указывает тип расщепления признака у особей второго поколения. Так, при трех доминантных генах А1; А2 и А3 и их рецессивных аллелях а1, а2, а3 во II поколении будут выявлены 64 варианта генотипов в соотношении 1 : 6: 15:20: 15:6: 1.
Если общая возможность развития признака связана с действием одного гена, то его принято называть главным (менделирующим) геном (олигогеном) и тогда признак наследуется в соответствии с законом Менделя. Полигены могут проявлять модифицирующее влияние на количественные признаки и составлять группу генов-модификаторов, то есть генов, которые, действуя каждый отдельно, проявляют слабое влияние на изменение в фенотипе, вызванное действием главного гена. Гены-модификаторы могут оказывать влияние и при отсутствии главного гена.
Полимерные гены способствуют увеличению изменчивости и формированию различных подгрупп (экотипов) внутри вида, так как они обеспечивают многообразные рекомбинации генотипов. Влияние рекомбинации и отбора в разных условиях среды способствует образованию несходных экотипов и повышению приспособленности вида к многообразию факторов внешней среды, формированию наследственной адаптации.
Гаметическая интеграция неаллельных генов. За последнее десятилетие расширились представления о наследовании и изменчивости признаков. Ранее полагали, что особенности потомства обусловлены свободной перекомбинацией генов как в процессе гаметогенеза, так и при оплодотворении. Однако многочисленные исследования показали, что изменчивость потомства, определяемая действием неаллельных генов, вызывается не только возможностью их случайной перекомбинации, а обусловлена определенной комбинацией гамет родителей. Таким образом, создается источник изменчивости и наследственности потомства за счет комбинаторики несходных гамет родителей. Несходство гамет по генетическому материалу может иметь место, как среди гамет самца, так и среди гамет самки. При комбинации несходных гамет родителей возникают новые межлокусные корреляции, происходит изменение частоты генов в данном локусе у потомства по сравнению с их частотой у исходных родительских форм. Создается новый тип связи между генетическими элементами у потомства, который получил название «гаметическая интеграция».
Гаметическая интеграция способствует формированию устойчивости разнообразных признаков и свойств, благоприятствующих повышению приспособленности, так как образуются адаптивные комплексы генов, которые повышают индивидуальную изменчивость организмов по степени их приспособленности к условиям среды. Гаметическая интеграция межлокусных корреляций обусловливает на фенотипическом уровне взаимодействие между локусэми неаллельных полигенов. Она может быть выявлена при помощи статистических параметров (σ, σ2, Cv).
Приспособленность организмов могут характеризовать величины среднего уровня жизнеспособности особей, их плодовитость, интенсивность развития всех особей данной группы (популяции).
Если в локусе одной из хромосом гаметы образовалась мутация, имеющая адаптивное значение, то при передаче ее в последующие поколения будет повышаться частота распространений мутации и одновременно на фоне гаметической интеграции будет увеличиваться частота других генов, входящих в хромосомы этой гаметы. Явление получило название «попутный транспорт генов».
Обобщение теории роли гаметической интеграции было сделано в трудах Майра, 1974; Левонтина, 1978; Животовского, 1984; и др.
Методы изучения изменчивости и наследуемости количественных признаков. Фенотипическую изменчивость количественных признаков определяют с помощью статистических параметров ( , σ, Cv), фенотипическую связь между признаками — применением r, b, rs и др. Для генетического анализа изменчивости количественных признаков требуется разложить фенотипическую изменчивость, выраженную через вариансу σр2, на составляющие вариансы: генотипическую (σg2) и паратипическую (σе2). Это позволяет установить в группе особей долю изменчивости, обусловленную их генетическим разнообразием, и долю изменчивости, связанную с влиянием факторов среды.
, σ, Cv), фенотипическую связь между признаками — применением r, b, rs и др. Для генетического анализа изменчивости количественных признаков требуется разложить фенотипическую изменчивость, выраженную через вариансу σр2, на составляющие вариансы: генотипическую (σg2) и паратипическую (σе2). Это позволяет установить в группе особей долю изменчивости, обусловленную их генетическим разнообразием, и долю изменчивости, связанную с влиянием факторов среды.
Теоретическая работа Фальконера (1985) показала, что популяционная средняя величина признака характеризует не только фенотипический его уровень, но и генотипический уровень в ряде поколений при сохранении факторов среды. Изменение среднего уровня признака при отборе происходит более эффективно в сторону его увеличения, чем в сторону уменьшения, что могло быть результатом действия «сильных» генов и материнского эффекта. Выяснено, что чем большее число локусов определяет уровень признака и чем длительнее отбор в ряде поколений, тем более выражена реакция на отбор.
Возникает вопрос: существуют ли тупики отбора, когда его действие прекращается? Оказывается, что постоянный процесс мутирования создает новый источник изменчивости и тупика отбора не возникает.
Для выявления наследуемости признака широко применяют методы корреляционного и дисперсионного анализа. Тем самым выявляется доля генетического влияния на признак в фенотипической изменчивости.
Наследуемость. В генетических исследованиях необходимо различать три близких, но разных понятия: наследственность, наследование и наследуемость. Наследственность — это биологическое свойство, проявляющееся в сходстве родителей и потомков. Наследование — это способ передачи наследственности родителей потомкам с помощью гамет и их хромосомного аппарата.
Наследуемость — это статистический термин, который применяют для обозначения доли общей фенотипической изменчивости, обусловленной генетическими факторами. Наследуемость характеризует количественный признак у группы животных и служит показателем для прогнозирования эффективности селекции по фенотипическим показателям признака.
Величина коэффициента наследуемости (h2) служит мерой выражения генетической детерминации изменчивости признака. Величина Л2 не может быть больше единицы и меньше нуля (то есть отрицательной). Формула для определения h2 выражена следующим образом:

Чем больше величина h2, тем больше изменчивость признака обусловлена генетическими факторами и тем меньше изменчивость, вызываемая факторами среды (σЕ2). При h2 менее 0,05 (то есть менее 5%) улучшение признака за счет массовой селекции малоэффективно. При h2>0,3 и не менее 0,7 селекция достаточно эффективна.
Особенности факторов среды и ряд характеристик популяции (направление отбора в ней, наличие инбридинга, миграции особей, интенсивность браковки в стаде и т. п.) влияют на величину коэффициента наследуемости. Большая изменчивость факторов среды уменьшает h2, а однородность условий увеличивает или стабилизирует его величину.
Вычисленные коэффициенты наследуемости могут характеризовать только данную популяцию и в данных условиях среды и отбора.
При рассмотрений величины h2 для разных признаков выявлена следующая тенденция: признаки, которые связаны с репродукцией и приспособленностью к процессу размножения, характеризуются невысоким коэффициентом наследуемости (плодовитость у свиней: h2 = 5%; яйценоскость у кур — 10%), а признаки, имеющие меньшее значение для приспособленности, характеризуются более высоким уровнем h2 (масса животных — 55—65%; толщина шпига у свиней —70%, молочность и жирномолочность — 35—40 %).
Определение коэффициента наследуемости с использованием коэффициентов корреляции (r) и регрессии (b). Метод коэффициентов путей, предложенный С. Райтом, позволяет выявить величину связи между фенотипической изменчивостью (следствие) и влиянием генотипа (причина) или среды (причина). Этот метод нашел применение для определения величины связи между генотипами родственных особей.
Для определения доли влияния генотипа на изменчивость признака используют коэффициенты путей между родственными животными, например между матерями и дочерьми или между полусибсами, отцами и сыновьями и т. п. Графическое изображение коэффициентов путей представлено на рис. 61,

где направление влияния причин указано стрелками. Из схемы следует, что генотип дочерей определяется генотипом матерей, величина влияния которого (а) равна 0,5. Остальная доля наследственности дочерей обусловлена генотипом отцов. Вместе с тем генотипы матерей оказывают влияние на их фенотипы (на схеме это обозначено h). Соответственно генотипы дочерей определяют их фенотип. Связь между фенотипами матерей и Дочерей может быть определена с помощью обычного коэффициента корреляции rмд. Из схемы видно, что звенья rмд, h, a, h образуют замкнутую цепь. В ней известны два звена:
и rмд, который вычисляют по конкретным данным. Так как связь между звеньями выражается произведением коэффициентов путей, то можно записать равенство: 0,5·h·h=rмд, или 0,5 h2 =rмд , тогда h2 =2rмд. При использовании Коэффициента регрессии bмд формула примет следующий вид: h2=2bмд. Полученная формула удвоенного коэффициента корреляций (или регрессии) между фенотипическими показателями матерей и их дочерей выражает величину коэффициента наследуемости h2, который определяет долю генетического влияния на фенотипическую изменчивость признака,
Если полученный коэффициент наследуемости — величина отрицательная или больше единицы, то причину этого можно объяснить следующим образом:
— группы животных, подвергнутых дисперсионному анализу, малочисленны, то есть объем выборки недостаточен;
— неоднозначное реагирование на изменений факторов среды материнских и дочерних особей и смена рангов тех и других. Нет смысла определять величину h2, если имеют место резкие различия в условиях между поколениями родителей и потомков:
— проявление межаллельного взаимодействий генов (доминирование, сверхдоминирование), влияние инбридинга.
Меньше подвержен воздействию «помех» коэффициент наследуемости, вычисленный с помощью регрессии: h2=2b. Это объясняется тем, что на величину коэффициента регрессии не влияет степень изменчивости признака. Если вычисление h2 ведется по потомству от одного отца, то регрессия дочерей по матерям в разрезе каждого производителя выражает величину аддитивной части наследуемости, то есть b = 0,5(σа2 : σр2).
Возможно также определение h2, основанное на сопоставлении признаков полусибсов, то есть у потомства от одного из родителей, по формуле h2=4rп/С или (реже) у полных сибсов: h2=2rc.
При сопоставлении результатов использования указанных формул выясняется, что они дают несколько различные величины h2 для одного и того же выборочного материала.
Для вычисления h2 можно применять дисперсионный анализ с разными типами статистических комплексов, используя формулу h2 =η2=Сx:Су (по ) или формулу Д. Снедекора: h2= σx2: (σx2+ σz2).
Коэффициент повторяемости. При проведении генетического анализа количественных признаков выявляют популяционный показатель, тесно связанный с h2 и имеющий важное значение для оценки наследственности. Этим показателем является коэффициент повторяемости (rw), Его свойства заключаются в следующем: a) rw — показатель генетического разнообразия; б) он является мерой верхнего предела коэффициента наследуемости; в) определяет надежность вносимых поправок в варьирующий признак с учетом изменения средовых факторов; г) служит мерой определения ошибки измеряемого признака.
При высокой величине повторяемости уровня признака особи сохраняют определенный его уровень, что дает возможность более эффективно вести селекцию.
Для вычисления rw чаще всего используют однофакторный дисперсионный комплекс. Коэффициент повторяемости находится в границах от 0 до +1. Для целей селекции предпочтительнее, если rw имеет большую величину, Считают, что если rw
Лекция № 17. Основные понятия генетики. Законы Менделя
Основные понятия генетики
Генетика — наука о закономерностях наследственности и изменчивости. Датой «рождения» генетики можно считать 1900 год, когда Г. Де Фриз в Голландии, К. Корренс в Германии и Э. Чермак в Австрии независимо друг от друга «переоткрыли» законы наследования признаков, установленные Г. Менделем еще в 1865 году.
Наследственность — свойство организмов передавать свои признаки от одного поколения к другому.
Изменчивость — свойство организмов приобретать новые по сравнению с родителями признаки. В широком смысле под изменчивостью понимают различия между особями одного вида.
Признак — любая особенность строения, любое свойство организма. Развитие признака зависит как от присутствия других генов, так и от условий среды, формирование признаков происходит в ходе индивидуального развития особей. Поэтому каждая отдельно взятая особь обладает набором признаков, характерных только для нее.
Фенотип — совокупность всех внешних и внутренних признаков организма.
Ген — функционально неделимая единица генетического материала, участок молекулы ДНК, кодирующий первичную структуру полипептида, молекулы транспортной или рибосомной РНК. В широком смысле ген — участок ДНК, определяющий возможность развития отдельного элементарного признака.
Генотип — совокупность генов организма.
Локус — местоположение гена в хромосоме.
Аллельные гены — гены, расположенные в идентичных локусах гомологичных хромосом.
Гомозигота — организм, имеющий аллельные гены одной молекулярной формы.
Гетерозигота — организм, имеющий аллельные гены разной молекулярной формы; в этом случае один из генов является доминантным, другой — рецессивным.
Рецессивный ген — аллель, определяющий развитие признака только в гомозиготном состоянии; такой признак будет называться рецессивным.
Доминантный ген — аллель, определяющий развитие признака не только в гомозиготном, но и в гетерозиготном состоянии; такой признак будет называться доминантным.
Методы генетики
Основным является гибридологический метод — система скрещиваний, позволяющая проследить закономерности наследования признаков в ряду поколений. Впервые разработан и использован Г. Менделем. Отличительные особенности метода: 1) целенаправленный подбор родителей, различающихся по одной, двум, трем и т. д. парам контрастных (альтернативных) стабильных признаков; 2) строгий количественный учет наследования признаков у гибридов; 3) индивидуальная оценка потомства от каждого родителя в ряду поколений.
Скрещивание, при котором анализируется наследование одной пары альтернативных признаков, называется моногибридным, двух пар — дигибридным, нескольких пар — полигибридным. Под альтернативными признаками понимаются различные значения какого-либо признака, например, признак — цвет горошин, альтернативные признаки — желтый цвет, зеленый цвет горошин.
Кроме гибридологического метода, в генетике используют: генеалогический — составление и анализ родословных; цитогенетический — изучение хромосом; близнецовый — изучение близнецов; популяционно-статистический метод — изучение генетической структуры популяций.
Генетическая символика
Предложена Г. Менделем, используется для записи результатов скрещиваний: Р — родители; F — потомство, число внизу или сразу после буквы указывает на порядковый номер поколения (F1 — гибриды первого поколения — прямые потомки родителей, F2 — гибриды второго поколения — возникают в результате скрещивания между собой гибридов F1); × — значок скрещивания; G — мужская особь; E — женская особь; A — доминантный ген, а — рецессивный ген; АА — гомозигота по доминанте, аа — гомозигота по рецессиву, Аа — гетерозигота.
Закон единообразия гибридов первого поколения, или первый закон Менделя
Другие материалы по теме:
Тема 2. Законы Менделя из книги «Генетика и селекция»
Успеху работы Менделя способствовал удачный выбор объекта для проведения скрещиваний — различные сорта гороха. Особенности гороха: 1) относительно просто выращивается и имеет короткий период развития; 2) имеет многочисленное потомство; 3) имеет большое количество хорошо заметных альтернативных признаков (окраска венчика — белая или красная; окраска семядолей — зеленая или желтая; форма семени — морщинистая или гладкая; окраска боба — желтая или зеленая; форма боба — округлая или с перетяжками; расположение цветков или плодов — по всей длине стебля или у его верхушки; высота стебля — длинный или короткий); 4) является самоопылителем, в результате чего имеет большое количество чистых линий, устойчиво сохраняющих свои признаки из поколения в поколение.
Опыты по скрещиванию разных сортов гороха Мендель проводил в течение восьми лет, начиная с 1854 года. 8 февраля 1865 года Г. Мендель выступил на заседании Брюннского общества естествоиспытателей с докладом «Опыты над растительными гибридами», где были обобщены результаты его работы.
Опыты Менделя были тщательно продуманы. Если его предшественники пытались изучить закономерности наследования сразу многих признаков, то Мендель свои исследования начал с изучения наследования всего лишь одной пары альтернативных признаков.
Мендель взял сорта гороха с желтыми и зелеными семенами и произвел их искусственное перекрестное опыление: у одного сорта удалил тычинки и опылил их пыльцой другого сорта. Гибриды первого поколения имели желтые семена. Аналогичная картина наблюдалась и при скрещиваниях, в которых изучалось наследование других признаков: при скрещивании растений, имеющих гладкую и морщинистую формы семян, все семена полученных гибридов были гладкими, от скрещивания красноцветковых растений с белоцветковыми все полученные — красноцветковые. Мендель пришел к выводу, что у гибридов первого поколения из каждой пары альтернативных признаков проявляется только один, а второй как бы исчезает. Проявляющийся у гибридов первого поколения признак Мендель назвал доминантным, а подавляемый — рецессивным.
( А — желтый цвет горошин, а — зеленый цвет горошин)
| Р | ♀ AA желтые | × | ♂ аа зеленые | ||
| Типы гамет |  А А | а | |||
| F1 | Aа желтые 100% | ||||
Закон расщепления, или второй закон Менделя
Г. Мендель дал возможность самоопылиться гибридам первого поколения. У полученных таким образом гибридов второго поколения проявился не только доминантный, но и рецессивный признак. Результаты опытов приведены в таблице.
| Признаки | Доминантные | Рецессивные | Всего | ||
|---|---|---|---|---|---|
| Число | % | Число | % | ||
| Форма семян | 5474 | 74,74 | 1850 | 25,26 | 7324 |
| Окраска семядолей | 6022 | 75,06 | 2001 | 24,94 | 8023 |
| Окраска семенной кожуры | 705 | 75,90 | 224 | 24,10 | 929 |
| Форма боба | 882 | 74,68 | 299 | 25,32 | 1181 |
| Окраска боба | 428 | 73,79 | 152 | 26,21 | 580 |
| Расположение цветков | 651 | 75,87 | 207 | 24,13 | 858 |
| Высота стебля | 787 | 73,96 | 277 | 26,04 | 1064 |
| Всего: | 14949 | 74,90 | 5010 | 25,10 | 19959 |
Анализ данных таблицы позволил сделать следующие выводы:
- единообразия гибридов во втором поколении не наблюдается: часть гибридов несет один (доминантный), часть — другой (рецессивный) признак из альтернативной пары;
- количество гибридов, несущих доминантный признак, приблизительно в три раза больше, чем гибридов, несущих рецессивный признак;
- рецессивный признак у гибридов первого поколения не исчезает, а лишь подавляется и проявляется во втором гибридном поколении.
Явление, при котором часть гибридов второго поколения несет доминантный признак, а часть — рецессивный, называют расщеплением. Причем, наблюдающееся у гибридов расщепление не случайное, а подчиняется определенным количественным закономерностям. На основе этого Мендель сделал еще один вывод: при скрещивании гибридов первого поколения в потомстве происходит расщепление признаков в определенном числовом соотношении.
Генетическая схема закона расщепления Менделя
( А — желтый цвет горошин, а — зеленый цвет горошин):
| P | ♀ Aa желтые | × | ♂ Aa желтые | ||
| Типы гамет | A | a | A | a | |
| F2 | AA желтые | Aa желтые 75% | Aa желтые | aa зеленые 25% | |
Закон чистоты гамет
Купить проверочные работы
и тесты по биологии




С 1854 года в течение восьми лет Мендель проводил опыты по скрещиванию растений гороха. Им было выявлено, что в результате скрещивания различных сортов гороха друг с другом гибриды первого поколения обладают одинаковым фенотипом, а у гибридов второго поколения имеет место расщепление признаков в определенных соотношениях. Для объяснения этого явления Мендель сделал ряд предположений, которые получили название «гипотезы чистоты гамет», или «закона чистоты гамет». Мендель предположил, что:
- за формирование признаков отвечают какие-то дискретные наследственные факторы;
- организмы содержат два фактора, определяющих развитие признака;
- при образовании гамет в каждую из них попадает только один из пары факторов;
- при слиянии мужской и женской гамет эти наследственные факторы не смешиваются (остаются чистыми).
В 1909 году В. Иогансен назовет эти наследственные факторы генами, а в 1912 году Т. Морган покажет, что они находятся в хромосомах.
Для доказательства своих предположений Г. Мендель использовал скрещивание, которое сейчас называют анализирующим (анализирующее скрещивание — скрещивание организма, имеющего неизвестный генотип, с организмом, гомозиготным по рецессиву). Наверное, Мендель рассуждал следующим образом: «Если мои предположения верны, то в результате скрещивания F1 с сортом, обладающим рецессивным признаком (зелеными горошинами), среди гибридов будут половина горошин зеленого цвета и половина горошин — желтого». Как видно из приведенной ниже генетической схемы, он действительно получил расщепление 1:1 и убедился в правильности своих предположений и выводов, но современниками он понят не был. Его доклад «Опыты над растительными гибридами», сделанный на заседании Брюннского общества естествоиспытателей, был встречен полным молчанием.
| Р | ♀ Аа желтые | × | ♂ aа зеленые |
| Типы гамет | A a | a | |
| F | Аа желтые 50% | аa зеленые 50% |
Цитологические основы первого и второго законов Менделя
Во времена Менделя строение и развитие половых клеток не было изучено, поэтому его гипотеза чистоты гамет является примером гениального предвидения, которое позже нашло научное подтверждение.
Явления доминирования и расщепления признаков, наблюдавшиеся Менделем, в настоящее время объясняются парностью хромосом, расхождением хромосом во время мейоза и объединением их во время оплодотворения. Обозначим ген, определяющий желтую окраску, буквой А , а зеленую — а . Поскольку Мендель работал с чистыми линиями, оба скрещиваемых организма — гомозиготны, то есть несут два одинаковых аллеля гена окраски семян (соответственно, АА и аа ). Во время мейоза число хромосом уменьшается в два раза, и в каждую гамету попадает только одна хромосома из пары. Так как гомологичные хромосомы несут одинаковые аллели, все гаметы одного организмы будут содержать хромосому с геном А , а другого — с геном а .
При оплодотворении мужская и женская гаметы сливаются, и их хромосомы объединяются в одной зиготе. Получившийся от скрещивания гибрид становится гетерозиготным, так как его клетки будут иметь генотип Аа ; один вариант генотипа даст один вариант фенотипа — желтый цвет горошин.
У гибридного организма, имеющего генотип Аа во время мейоза, хромосомы расходятся в разные клетки и образуется два типа гамет — половина гамет будет нести ген А , другая половина — ген а . Оплодотворение — процесс случайный и равновероятный, то есть любой сперматозоид может оплодотворить любую яйцеклетку. Поскольку образовалось два типа сперматозоидов и два типа яйцеклеток, возможно возникновение четырех вариантов зигот. Половина из них — гетерозиготы (несут гены А и а ), 1/4 — гомозиготы по доминантному признаку (несут два гена А ) и 1/4 — гомозиготы по рецессивному признаку (несут два гена а ). Гомозиготы по доминанте и гетерозиготы дадут горошины желтого цвета (3/4), гомозиготы по рецессиву — зеленого (1/4).
Закон независимого комбинирования (наследования) признаков, или третий закон Менделя
Организмы отличаются друг от друга по многим признакам. Поэтому, установив закономерности наследования одной пары признаков, Г. Мендель перешел к изучению наследования двух (и более) пар альтернативных признаков. Для дигибридного скрещивания Мендель брал гомозиготные растения гороха, отличающиеся по окраске семян (желтые и зеленые) и форме семян (гладкие и морщинистые). Желтая окраска ( А ) и гладкая форма ( В ) семян — доминантные признаки, зеленая окраска ( а ) и морщинистая форма ( b ) — рецессивные признаки.
Скрещивая растение с желтыми и гладкими семенами с растением с зелеными и морщинистыми семенами, Мендель получил единообразное гибридное поколение F1 с желтыми и гладкими семенами. От самоопыления 15-ти гибридов первого поколения было получено 556 семян, из них 315 желтых гладких, 101 желтое морщинистое, 108 зеленых гладких и 32 зеленых морщинистых (расщепление 9:3:3:1).
Анализируя полученное потомство, Мендель обратил внимание на то, что: 1) наряду с сочетаниями признаков исходных сортов (желтые гладкие и зеленые морщинистые семена), при дигибридном скрещивании появляются и новые сочетания признаков (желтые морщинистые и зеленые гладкие семена); 2) расщепление по каждому отдельно взятому признаку соответствует расщеплению при моногибридном скрещивании. Из 556 семян 423 были гладкими и 133 морщинистыми (соотношение 3:1), 416 семян имели желтую окраску, а 140 — зеленую (соотношение 3:1). Мендель пришел к выводу, что расщепление по одной паре признаков не связано с расщеплением по другой паре. Для семян гибридов характерны не только сочетания признаков родительских растений (желтые гладкие семена и зеленые морщинистые семена), но и возникновение новых комбинаций признаков (желтые морщинистые семена и зеленые гладкие семена).
| Р | ♀ АABB желтые, гладкие | × | ♂ aаbb зеленые, морщинистые |
| Типы гамет |  AB AB | ab | |
| F1 | AaBb желтые, гладкие, 100% | ||
| P | ♀ АaBb желтые, гладкие | × | ♂ AаBb желтые, гладкие |
| Типы гамет | AB Ab aB ab | AB Ab aB ab | |
Генетическая схема закона независимого комбинирования признаков:
| Гаметы: | ♂ | AB | Ab | aB | ab |
| ♀ | |||||
| AB | AABB желтые гладкие | AABb желтые гладкие | AaBB желтые гладкие | AaBb желтые гладкие | |
| Ab | AABb желтые гладкие | AАbb желтые морщинистые | AaBb желтые гладкие | Aabb желтые морщинистые | |
| aB | AaBB желтые гладкие | AaBb желтые гладкие | aaBB зеленые гладкие | aaBb зеленые гладкие | |
| ab | AaBb желтые гладкие | Aabb желтые морщинистые | aaBb зеленые гладкие | aabb зеленые морщинистые | |
Анализ результатов скрещивания по фенотипу: желтые, гладкие — 9/16, желтые, морщинистые — 3/16, зеленые, гладкие — 3/16, зеленые, морщинистые — 1/16. Расщепление по фенотипу 9:3:3:1.
Анализ результатов скрещивания по генотипу: AaBb — 4/16, AABb — 2/16, AaBB — 2/16, Aabb — 2/16, aaBb — 2/16, ААBB — 1/16, Aabb — 1/16, aaBB — 1/16, aabb — 1/16. Расщепление по генотипу 4:2:2:2:2:1:1:1:1.
Если при моногибридном скрещивании родительские организмы отличаются по одной паре признаков (желтые и зеленые семена) и дают во втором поколении два фенотипа (2 1 ) в соотношении (3 + 1) 1 , то при дигибридном они отличаются по двум парам признаков и дают во втором поколении четыре фенотипа (2 2 ) в соотношении (3 + 1) 2 . Легко посчитать, сколько фенотипов и в каком соотношении будет образовываться во втором поколении при тригибридном скрещивании: восемь фенотипов (2 3 ) в соотношении (3 + 1) 3 .
Если расщепление по генотипу в F2 при моногибридном поколении было 1:2:1, то есть было три разных генотипа (3 1 ), то при дигибридном образуется 9 разных генотипов — 3 2 , при тригибридном скрещивании образуется 3 3 — 27 разных генотипов.
Третий закон Менделя справедлив только для тех случаев, когда гены анализируемых признаков находятся в разных парах гомологичных хромосом.
Цитологические основы третьего закона Менделя
Пусть А — ген, обусловливающий развитие желтой окраски семян, а — зеленой окраски, В — гладкая форма семени, b — морщинистая. Скрещиваются гибриды первого поколения, имеющие генотип АаВb . При образовании гамет из каждой пары аллельных генов в гамету попадает только один, при этом в результате случайного расхождения хромосом в первом делении мейоза ген А может попасть в одну гамету с геном В или с геном b , а ген а — с геном В или с геном b . Таким образом, каждый организм образует четыре сорта гамет в одинаковом количестве (по 25%): АВ , Ab , aB , ab . Во время оплодотворения каждый из четырех типов сперматозоидов может оплодотворить любую из четырех типов яйцеклеток. В результате оплодотворения возможно появление девяти генотипических классов, которые дадут четыре фенотипических класса.
Перейти к лекции №16 «Онтогенез многоклеточных животных, размножающихся половым способом»
Перейти к лекции №18 «Сцепленное наследование»
Смотреть оглавление (лекции №1-25)