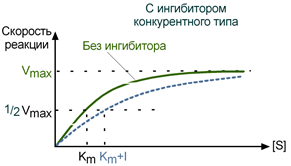
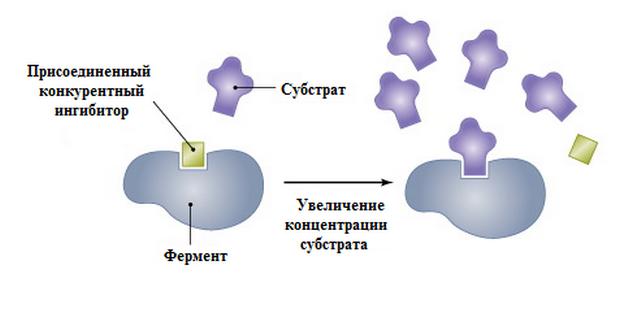
При таком виде ингибирования ингибитор конкурирует с субстратом за активный центр (за контактный участок), что приводит к уменьшению связывания субстрата с ферментом и снижению катализа. Используя терминологию кинетики Михаэлиса-Ментен можно сказать, что конкурентный ингибитор уменьшает сродство фермента к субстрату, повышая константу Михаэлиса (Km), максимальная скорость реакции (Vmax) остается при этом неизменнной.
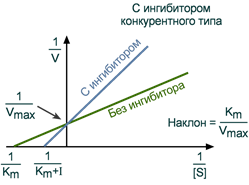
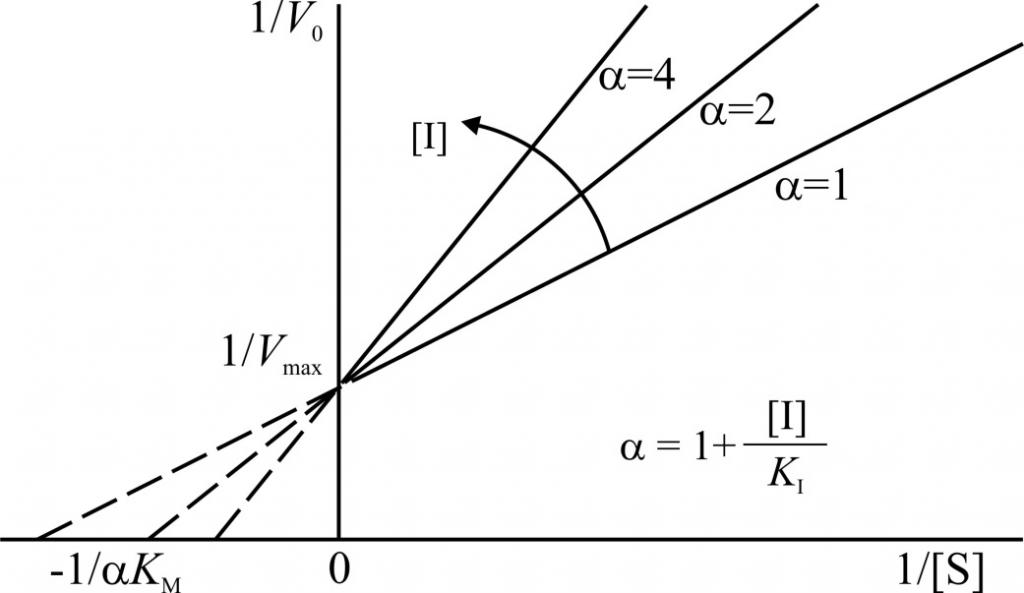
Зависимость в координатах Лайнуивера-Берка имеет вид пучка прямых, пересекающихся на оси ординат. Конкурентный ингибитор увеличивает Кm и не изменяет Vmax.
График конкурентного ингибирования в координатах Михаэлиса-Ментен
График конкурентного ингибирования в координатах Лайнуивера-Берка
Кинетика неконкурентного ингибирования
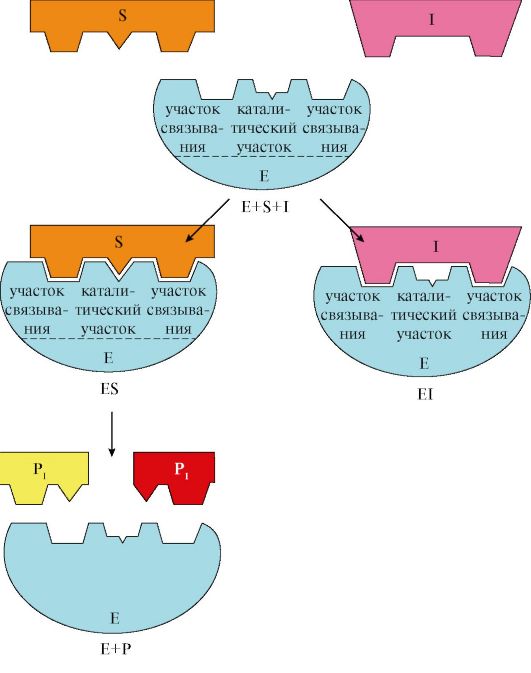
Особенностью неконкурентного ингибитора является его способность связываться с ферментом не в активном центре, и изменение концентрации субстрата никак не влияет на это связывание. В то же время неконкурентный ингибитор не мешает связыванию субстрата с активным центром. В результате формируется тройной комплекс фермент-субстрат-ингибитор (E-S-I), в котором фермент уже не способен изменить свою конформацию и обеспечить проведение реакции. Количество «работоспособных» комплексов E-S при этом снижается.
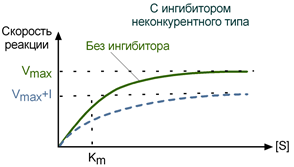
Максимальная скорость реакции (Vmax) при неконкурентном ингибировании снижается, константа Михаэлиса (Km) не изменяется, т.е. добавление дополнительного субстрата не может повлиять на состояние активного центра и работу фермента.
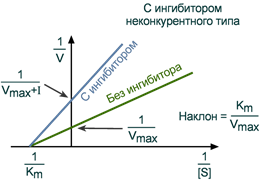
Зависимость в координатах Лайнуивера-Берка имеет вид пучка прямых, пересекающихся на оси абсцисс. Неконкурентный ингибитор не изменяет Кm и снижает Vmax.
График неконкурентного ингибирования в координатах Михаэлиса-Ментен
График неконкурентного ингибирования в координатах Лайнуивера-Берка
Кинетика бесконкурентного ингибирования
Бесконкурентный ингибитор способен связываться только с уже образовавшимся комплексом E-S, но не со свободным ферментом. Связывание происходит либо на молекуле субстрата, либо с ферментом, уже начавшем катализ и изменившим свою конформацию. Образовавшийся тройной комплекс E-S-I также непродуктивен.
Максимальная скорость реакции (Vmax) и константа Михаэлиса (Km) при бесконкурентном ингибировании снижаются. Добавление дополнительного субстрата не может повлиять на состояние активного центра и работу фермента.
Конкурентное ингибирование: определение, особенности и примеры
Все биохимические реакции, протекающие в организме, подвержены специфическому контролю, который осуществляется через активирующее или ингибирующее воздействие на регуляторные ферменты. Последние обычно находятся в начале цепочек метаболических превращений и либо запускают многоэтапный процесс, либо тормозят его. Регулированию также подвергаются некоторые единичные реакции. Конкурентное ингибирование является одним из основных механизмов контроля каталитической активности ферментов.
Что такое ингибирование?
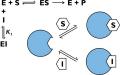
Механизм ферментативного катализа основан на связывании активного центра энзима с молекулой субстрата (комплекс ES), в результате чего происходит химическая реакция с образованием и освобождением продукта (E+S = ES = EP = E+P).
Ингибированием фермента называют снижение скорости или полную остановку процесса катализа. В более узком смысле под этим термином подразумевают уменьшение сродства активного центра к субстрату, что достигается путем связывания молекул энзимов с веществами-ингибиторами. Последние могут действовать различными способами, на основании чего поделены на несколько типов, которым соответствуют одноименные механизмы ингибирования.
Основные типы ингибирования
По характеру протекания процесса ингибирование бывает двух видов:
Необратимое – вызывает стойкие изменения в молекуле фермента, лишающие ее функциональной активности (последняя не подлежит восстановлению). Оно может иметь как специфический, так и неспецифический характер. Ингибитор прочно связывается с энзимом путем ковалентного взаимодействия.
Обратимое – основной вид негативной регуляции ферментов. Осуществляется за счет обратимого специфического присоединения ингибитора к белку-энзиму слабыми нековалентными связями, поддается кинетическому описанию по уравнению Михаэлиса-Ментен (исключение составляет аллостерическая регуляция).
Выделяют два основных типа обратимого ингибирования ферментов: конкурентное (может быть ослаблено увеличением концентрации субстрата) и неконкурентное. В последнем случае происходит снижение максимально возможной скорости катализа.
Основная разница между конкурентным и неконкурентным ингибированием заключается в месте присоединения регуляторного вещества к ферменту. В первом случае ингибитор связывается непосредственно с активным центром, а во втором – с другим участком энзима, либо с фермент-субстратным комплексом.
Существует также смешанный тип ингибирования, при котором связывание с ингибитором не предотвращает образование ES, но замедляет катализ. В этом случае вещество-регулятор находится в составе двойных или тройных комплексов (EI и EIS). При бесконкурентном типе фермент связывается только с ES.
Особенности обратимого конкурентного ингибирования ферментов
Конкурентный механизм ингибирования основан на структурном сходстве регуляторного вещества с субстратом. В результате образуется комплекс активного центра с ингибитором, условно обозначаемый как EI.
Обратимое конкурентное ингибирование имеет следующие особенности:
связывание с ингибитором происходит в активном центре;
инактивация молекулы фермента обратима;
ингибирующий эффект может быть уменьшен увеличением концентрации субстрата;
ингибитор не влияет на максимальную скорость ферментативного катализа;
комплекс EI может распадаться, что характеризуется соответствующей константой диссоциации.
При таком типе регуляции ингибитор и субстрат как бы соперничают (конкурируют) друг с другом за место в активном центре, откуда и произошло название процесса.
В итоге конкурентное ингибирование можно определить как обратимый процесс торможения ферментативного катализа, основанный на специфическом сродстве активного центра к веществу-ингибитору.
Механизм действия
Связывание ингибитора с активным центром препятствует образованию фермент-субстратного комплекса, необходимого для осуществления катализа. В итоге молекула энзима становится неактивной. Тем не менее каталитический центр может связаться не только с ингибитором, но и с субстратом. Вероятность образования того или иного комплекса зависит от соотношения концентраций. Если молекул субстрата значительно больше, то фермент будет реагировать с ними чаще, чем с ингибитором.
Влияние на скорость химической реакции
Степень торможения катализа при конкурентном ингибировании определяется тем, какое количество фермента будут образовывать EI-комплексы. При этом можно увеличить концентрацию субстрата до такой степени, что роль ингибитора будет вытеснена, а скорость катализа достигнет максимально возможного значения, соответствующего величине Vmax по уравнению Михаэлиса-Ментен.
Такое явление объясняется сильным разбавлением ингибитора. Как следствие, вероятность связывания с ним молекул фермента сводится к нулю, а активные центры реагируют только с субстратом.
Кинетические зависимости ферментативной реакции при участии конкурентного ингибитора
Конкурентное ингибирование увеличивает константу Михаэлиса (Km), которая равна концентрации субстрата, необходимой для достижения ½ максимальной скорости катализа в начале реакции. Количество фермента, гипотетически способного связаться с субстратом, остается постоянным, а число фактически образующихся ES-комплексов зависит только от концентрации последнего (комплексы EI не постоянны и могут быть вытеснены субстратом).
Конкурентное ингибирование ферментов легко определить по графикам кинетической зависимости, построенным для разных концентраций субстрата. В этом случае величина Km будет меняться, а Vmax оставаться постоянной величиной.
При неконкурентном ингибировании все наоборот: ингибитор связывается вне активного центра и присутствие субстрата никак не может на это повлиять. В результате часть молекул фермента «выключается» из катализа, и максимально возможная скорость снижается. Тем не менее активные молекулы энзима могут беспрепятственно связываться с субстратом как при маленькой, так и при высокой концентрации последнего. Следовательно, константа Михаэлиса остается постоянной.
Графики конкурентного ингибирования в системе двойных обратных координат представляют собой несколько прямых, пересекающих ось ординат в точке 1/Vmax. Каждая прямая соответствует определенной концентрации субстрата. Разные точки пересечения с осью абсцисс (1/[S]) говорят об изменении константы Михаэлиса.
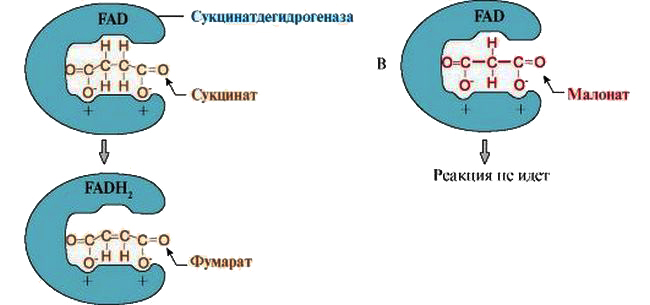
Действие конкурентного ингибитора на примере малоната
Типичным примером конкурентного ингибирования является процесс снижения активности сукцинатдегидрогиназы — фермента, катализирующего окисление янтарной кислоты (сукцината) в фумаровую. В роли ингибитора здесь выступает малонат, имеющий структурное сходство с сукцинатом.
Добавление ингибитора в среду вызывает образование комплексов малоната с сукцинатдегидрогиназой. Такая связь не вызывает повреждения активного центра, но блокирует его доступность для янтарной кислоты. Увеличение концентрации сукцината снижает ингибирующий эффект.
Использование в медицине
На механизме конкурентного ингибирования основано действие многих лекарственных препаратов, представляющих собой структурные аналоги субстратов некоторых метаболических путей, торможение которых является необходимой частью лечения заболеваний.
Например, для улучшения проводимости нервных импульсов при мышечных дистрофиях требуется повысить уровень ацетилхолина. Это достигается угнетением активности гидролизующей его ацетилхолинэстеразы. В роли ингибиторов выступают четвертичные аммониевые основания, входящие в состав лекарственных препаратов (прорезин, эндрофоний и т. д.).
В особую группу выделяют антиметаболиты, которые помимо ингибирующего действия проявляют свойства псевдосубстрата. В таком случае формирование комплекса EI приводит к образованию биологически инертного аномального продукта. К антиметаболитам относят сульфаниламиды (используются при лечении бактериальных инфекций), аналоги нуклеотидов (применяются для остановки клеточного роста раковой опухоли) и т. д.
Кинетика и термодинамика ферментативных реакций (стр. 4 )
Из за большого объема этот материал размещен на нескольких страницах: 1 2 3 4 5 6 7 8 9
Смешанное конкурентное-неконкурентное ингибирование. В этом варианте ингибирования Ki2˃ Ki1. Сродство фермента к субстрату в присутствии ингибитора данного типа увеличивается, а максимальная скорость ферментативной реакции снижается (рис. 2.8).
, (8)
Рис. 2.8. Смешанное конкурентное-неконкурентное
Смешанное неконкурентное-бесконкурентное ингибирование. В этом случае Ki1 ˃ Ki2. Этот ингибитор снижает и константу Михаэлиса и максимальную скорость реакции (рис. 2.9).
Примером смешанного ингибирования является воздействие ртутьорганического соединения мертиолата на сахаразу грибов. Это вещество широко используется в промышленности для подавления роста микромицетов.
2.5. Методы определения константы ингибирования
Величины Кi можно измерить различными методами, как экспериментально, так и при помощи расчетов.
Если известна только одна концентрация ингибитора, то константу ингибирования можно определить, используя графики в координатах Лайнуивера-Берка. Найдя кажущиеся и истинные значения константы Михаэлиса и максимальной скорости для двух графиков (в отсутствии и в присутствии ингибитора) и подставив эти значения в формулы (2), (4), (7), (9), (10), рассчитывают ингибиторную константу.
Наиболее удобен для определения константы ингибирования метод Диксона. Этот простой графический метод позволяет определять Кi непосредственно. Если определять скорость реакции в условиях постоянной концентрации субстрата и различных концентраций ингибитора, то график зависимости 1/v от [I] будет представлять собой прямую (рис. 2.10.). В этом случае для определения ингибиторной константы достаточно определить скорость реакции всего для двух концентраций субстрата.
Рис. 2.10. Графическое определение ингибиторных констант (метод Диксона). Во всех случаях S1˃S2
2.6. Субстратное ингибирование
Для многих ферментативных реакций при увеличении концентрации субстрата начальная скорость ферментативной реакции проходит через максимум, а затем уменьшается. Подобного рода зависимость можно описать, исходя из предположения об образовании в процессе реакции непродуктивного тройного комплекса ЕS2. В этом случае кинетическая схема ферментативной реакции выглядит следующим образом:
Зависимость скорости реакции от концентрации субстрата после математической обработки этой схемы имеет вид:





 ,
,  (8)
(8)
 ,
,  (9)
(9)


 (11)
(11)