- Окисление жирных кислот (β-окисление)
- Этапы окисления жирных кислот
- Расчет энергетического баланса β-окисления
- B-Окисление жирных кислот: реакции одного оборота цикла, регуляция, энергетический баланс окисления стеариновой и олеиновой кислот (сравнить).
- Бета окисление стеариновой кислоты уравнение реакции
- 🎥 Видео
Видео:Биохимия | Бета-окисление жирных кислот: насыщенных, ненасыщенных, с нечетным количеством атомов ССкачать

Окисление жирных кислот (β-окисление)
Для преобразования энергии, заключенной в жирных кислотах, в энергию связей АТФ существует метаболический путь окисления жирных кислот до СО2 и воды, тесно связанный с циклом трикарбоновых кислот и дыхательной цепью. Этот путь называется β-окисление, т.к. происходит окисление 3-го углеродного атома жирной кислоты (β-положение) в карбоксильную группу, одновременно от кислоты отщепляется ацетильная группа, включающая С 1 и С 2 исходной жирной кислоты.
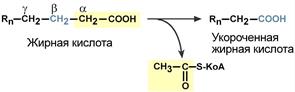
Элементарная схема β-окисления
Реакции β-окисления происходят в митохондриях большинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из крови или появляющиеся при липолизе собственных внутриклеточных ТАГ. Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом:
Пальмитоил-SКоА + 7ФАД + 7НАД + + 7Н2O + 7HS-KoA → 8Ацетил-SКоА + 7ФАДН2 + 7НАДН
Этапы окисления жирных кислот
1. Прежде, чем проникнуть в матрикс митохондрий и окислиться, жирная кислота должна активироваться в цитозоле. Это осуществляется присоединением к ней коэнзима А с образованием ацил-SКоА. Ацил-SКоА является высокоэнергетическим соединением. Необратимость реакции достигается гидролизом дифосфата на две молекулы фосфорной кислоты.
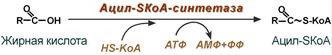
Реакция активации жирной кислоты
2. Ацил-SКоА не способен проходить через митохондриальную мембрану, поэтому существует способ переноса жирной кислоты в комплексе с витаминоподобным веществом карнитином (витамин В11). На наружной мембране митохондрий имеется фермент карнитин-ацилтрансфераза I.
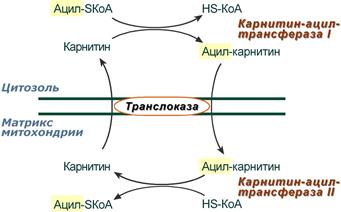
Карнитин-зависимый транспорт жирных кислот в митохондрию
3. После связывания с карнитином жирная кислота переносится через внутреннюю митохондриальную мембрану транслоказой . На внутренней стороне этой мембраны фермент карнитин-ацилтрансфераза II вновь образует ацил-SКоА, который вступает на путь β-окисления.
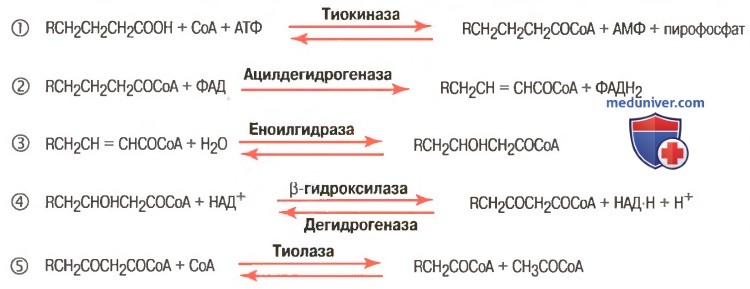
4. Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление (ацил-SКоА-дегидрогеназа), гидратирование (еноил-SКоА-гидратаза) и вновь окисление 3-го атома углерода (гидроксиацил-SКоА-дегидрогеназа). В последней, трансферазной, реакции от жирной кислоты отщепляется ацетил-SКоА . К оставшейся (укороченной на два углерода) жирной кислоте присоединяется HS-КоА, и она возвращается к первой реакции. Все повторяется до тех пор, пока в последнем цикле не образуются два ацетил-SКоА.
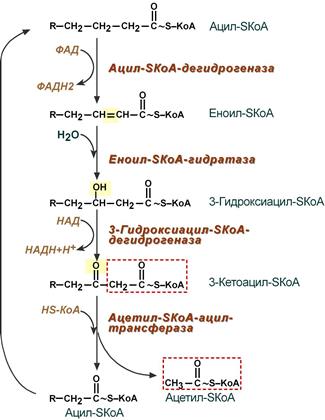
Последовательность реакций β-окисления жирных кислот
Расчет энергетического баланса β-окисления
Ранее при расчете эффективности окисления коэффициент P/O для НАДH принимался равным 3,0, для ФАДH2 – 2,0.
По современным данным значение коэффициента P/O для НАДH соответствует 2,5, для ФАДH2 – 1,5.
При расчете количества АТФ, образуемого при β-окислении жирных кислот необходимо учитывать:
- количество образуемого ацетил-SКоА – определяется обычным делением числа атомов углерода в жирной кислоте на 2.
- число циклов β-окисления . Число циклов β-окисления легко определить исходя из представления о жирной кислоте как о цепочке двухуглеродных звеньев. Число разрывов между звеньями соответствует числу циклов β-окисления. Эту же величину можно подсчитать по формуле (n/2 -1), где n – число атомов углерода в кислоте.
- число двойных связей в жирной кислоте. В первой реакции β-окисления происходит образование двойной связи при участии ФАД. Если двойная связь в жирной кислоте уже имеется, то необходимость в этой реакции отпадает и ФАДН2 не образуется. Количество недополученных ФАДН2 соответствует числу двойных связей. Остальные реакции цикла идут без изменений.
- количество энергии АТФ , потраченной на активацию (всегда соответствует двум макроэргическим связям).
Видео:Биохимия: Бета окислениеСкачать

B-Окисление жирных кислот: реакции одного оборота цикла, регуляция, энергетический баланс окисления стеариновой и олеиновой кислот (сравнить).
Реакции β-окисление ЖК
1. β-окисление начинается с дегидрирования ацил-КоА ФАД-зависимой Ацил-КоА дегидрогеназой с образованием двойной связи (транс) между α- и β-атомами С в Еноил-КоА. Восстановленный ФАДН2 окисляясь в ЦПЭ, обеспечивает синтез 2 молекул АТФ;
2. Еноил-КоА гидратаза присоединяет воду к двойной связи Еноил-КоА с образованием β-оксиацил-КоА;
3. β-оксиацил-КоА окисляется НАД зависимой дегидрогеназой до β-кетоацил-КоА. Восстановленный НАДН2, окисляясь в ЦПЭ, обеспечивает синтез 3 молекул АТФ;
4. Тиолаза с участием HКоА отщепляет от β-кетоацил-КоА Ацетил-КоА. В результате 4 реакций образуется Ацил-КоА, который короче предыдущего Ацил-КоА на 2 углерода. Образованный Ацетил-КоА окисляясь в ЦТК, обеспечивает синтез в ЦПЭ 12 молекул АТФ.
Затем Ацил-КоА снова вступает в реакции β-окисления. Циклы продолжаются до тех пор, пока Ацил-КоА не превратится в Ацетил-КоА с 2 атома С (если ЖК имела четное количество атомов С) или Бутирил-КоА с 3 атомами С (если ЖК имела нечетное количество атомов С).

Регуляция скорости β-окисления ЖК
β-окисление активируют: НАД + , АДФ (энергодефицит), ЖК, глюкагон, адреналин.
β-окисление ингибируют: НАДH2, АТФ, инсулин.
Голод, физическая нагрузка → ↑ глюкагон, ↑ адреналин → липолиз ТГ в адипоцитах → ↑ ЖК в крови → ↑ β-окисление в аэробных условиях в мышцах, печени → 1) ↑АТФ; 2) ↑АТФ, ↑НАДH2, ↑Ацетил-КоА, (↑ЖК) → ↓ гликолиз → ↑ экономию глюкозы, необходимую для нервной ткани, эритроцитов и т.д.
Пища → ↑ инсулин → ↑ гликолиз → ↑ Ацетил-КоА → ↑ синтез малонил-КоА и ЖК
↑ синтез малонил-КоА → ↑ малонил-КоА → ↓ карнитинацилтрансферазы I в печени → ↓ транспорт ЖК в матрикс митохондрий → ↓ ЖК в матриксе → ↓ β-окисление ЖК
Энергетический баланс окисления насыщенных ЖК
С четным количеством атомов углерода
При активации ЖК затрачивается 2 макроэргической связи АТФ.
При окислении насыщенной ЖК с четным количеством атомов С образуются только ФАДН2, НАДН2 и Ацетил-КоА.
За 1 цикл β-окисления образуется 1 ФАДН2, 1 НАДН2 и 1 Ацетил-КоА, которые при окислении дают 2+3+12=17 АТФ.
Количество циклов при β-окислении ЖК = количество атомов С в (ЖК/2)-1. Стеариновая кислота при β-окислении проходит (18/2)-1 = 8 циклов. За 8 циклов образуется 17*8=136 АТФ.
Последний цикл β-окисления сопровождается образованием дополнительной Ацетил-КоА, которая при окислении дает 12 АТФ.
Таким образом, при окислении стеариновой кислоты образуется: -2+136+12=146 АТФ.
Энергетический баланс окисления ненасыщенных ЖК
С четным количеством атомов углерода
Около половины ЖК в организме человека ненасыщенные. β-окисление этих кислот идёт обычным путём до тех пор, пока двойная связь не окажется между 3 и 4 атомами С. Затем фермент еноил-КоА изомераза перемещает двойную связь из положения 3-4 в положение 2-3 и изменяет цис-конформацию двойной связи на транс-, которая необходима для β-окисления. В этом цикле β-окисления, так как двойная связь в ЖК уже имеется, первая реакция дегидрирования не происходит и ФАДН2 не образуется. Далее циклы β-окисления продолжаются, не отличаясь от обычного пути.
Энергетический баланс рассчитывается также как и для насыщенных ЖК с четным количеством атомов С, только на каждую двойную связь недосчитывают 1 ФАДН2 и соответственно 2 АТФ.
Суммарное уравнение β-окисления пальмитолеил-КоА:
С15Н29СО-КоА + 6 ФАД + 7 НАД + + 7 HSKoA → 8 CH3-CO-KoA + 6 ФАДН2 + 7 НАДН2
?Энергетический баланс β-окисления олеиновой кислоты: -2+8*12+7*2+8*3=132 АТФ.
Биохимия
25. Окисление глицерина до Н2О и СО2: схема, энергетический баланс.
1. Окисление глицерина начинается с его фосфорилирования и превращения в глицеринофосфорную к-ту, к-рая подвергается окислению с образованием фосфодиоксиацетона, переходящего в фосфоглицериновый альдегид. Его последующие превращения идут по схеме гликолиза, а затем приводят к образованию двууглеродистого компонента— ацетил-КоА. Активированные высшие жирные к-ты в виде соединений с КоА реагируют с карнитином, образуя его производные, способные проникать через мембрану митохондрий. Внутри митохондрий жирные к-ты подвергаются последовательному β-окислению с освобождением ацетил-КоА, к-рый используется в цикле трикарбоновых кислот или в реакциях биосинтеза. Ж. о. регулируется ц. н. с., гормонами гипофиза, надпочечников, половых желез и др
2.Окисление глицерина в тканях:
— реакции вовлечения глицерина в гликолиз (образование диоксиацетонфосфата, а затем 3-фосфоглицеринового альдегида- метаболита гликолиза).
— энергетический эффект окисления глицерина в аэробных условиях (22 молекулы АТФ).
глицерин(-АТФ)->фосфоглицерин(+НАДН)- >3-фосфоглицериновый альдегид->. (+2АТФ, +НАДН)->пируват(+НАДН)->ацетилкофермент А->СО2+Н2О+12АТФ (в цикле Кребса).
+ это значит образуется, — расходуется (обозначения мои).
Из одного НАДН получается 3АТФ в дыхательной цепи.
Итого 3НАДН+2АТФ+12АТФ-1АТФ=9АТФ+13АТФ=22АТФ
26. Окисление ТГ до Н2О и СО2: схема, энергетический баланс.
Липолиз. Липолиз в жировой ткани активируется при дефиците глюкозы в крови (постабсорбционный период, голодание, физическая нагрузка). Процесс стимулируется глюкагоном, адреналином, в меньшей степени СТГ и глюкокортикоидами.
В результате липолиза концентрация свободных жирных кислот в крови возрастает в 2 раза.
Видео:β-окисление жирных кислотСкачать

Бета окисление стеариновой кислоты уравнение реакции
а) Расщепление жирных кислот до ацетилкоэнзима А путем бета-окисления. Молекула жирной кислоты расщепляется в митохондрии путем постепенного отщепления двууглеродных фрагментов в виде ацетилкоэнзима А (ацетил-КоА). Этот процесс, ход которого представлен на рисунке ниже, называют процессом бета-окисления.
 Бета-окисление жирных кислот в результате дает ацетилкоэнзим А
Бета-окисление жирных кислот в результате дает ацетилкоэнзим А
Обратите внимание, что первый этап бета-окисления представляет собой взаимодействие молекулы жирной кислоты с коэнзимом А (КоА) с образованием ацил-КоА жирной кислоты. В уравнениях 2, 3 и 4 бета-углерод (второй углерод справа) ацил-КоА жирной кислоты взаимодействует с молекулой кислорода, вследствие этого бета-углерод окисляется.
В правой части уравнения 5 двууглеродная часть молекулы отщепляется, образуя ацетил-КоА, выделяющийся во внеклеточную жидкость. В то же время другая молекула КоА взаимодействует с концом оставшейся части молекулы жирной кислоты, вновь формируя ацил-КоА жирной кислоты. Сама молекула жирной кислоты в это время становится короче на 2 атома углерода, т.к. первый ацетил-КоА уже отделился от ее терминали.
Затем эта укоротившаяся молекула ацил-КоА жирной кислоты выделяет еще 1 молекулу ацетил-КоА, что приводит к укорочению исходной молекулы жирной кислоты еще на 2 атома углерода. Кроме высвобождения молекул ацетил-КоА из молекул жирных кислот в ходе этого процесса выделяются 4 атома углерода.
б) Окисление ацетил-КоА. Образующиеся в митохондриях в ходе процесса бета-окисления жирных кислот молекулы ацетил-КоА немедленно поступают в цикл лимонной кислоты и, взаимодействуя прежде всего с щавелево-уксусной кислотой, образуют лимонную кислоту, которая затем последовательно окисляется посредством хемоосмотическои. системы окисления митохондрий. Чистый выход реакции цикла лимонной кислоты на 1 молекулу аце-тил-КоА составляет:

Таким образом, после начального расщепления жирной кислоты с образованием ацетил-КоА окончательное их расщепление осуществляется так же, как расщепление ацетил-КоА, образовавшегося из пировиноградной кислоты в процессе метаболизма глюкозы. Образующиеся при этом атомы водорода окисляются той же системой окисления митохондрий, которая используется в процессе окисления углеводов, с образованием большого количества аденозинтрифосфата.
в) При окислении жирных кислот образуется огромное количество АТФ. На рисунке выше показано, что 4 атома водорода, высвобождающиеся при отделении ацетил-КоА от цепочки жирной кислоты, выделяются в виде ФАДН2, НАД-Н и Н+, поэтому при расщеплении 1 молекулы стеариновой кислоты образуется, кроме 9 молекул ацетил-КоА, еще 32 атома водорода. В процессе расщепления каждой из 9 молекул ацетил-КоА в цикле лимонной кислоты выделяются еще 8 атомов водорода, что в итоге дает 72 атома водорода.
Суммарно при расщеплении 1 молекулы стеариновой кислоты выделяются 104 атома водорода. Из этого общего количества 34 атома выделяются, будучи связанными с флавопротеинами, а остальные 70 высвобождаются в форме, связанной с никотинамидадениндинуклеотидом, т.е. в виде НАД-Н+ и Н+.
Окисление водорода, связанного с этими двумя типами веществ, осуществляется в митохондриях, но они вступают в процесс окисления в разных точках, поэтому окисление каждого из 34 атомов водорода, связанных с флавопротеинами, приводит к выделению 1 молекулы АТФ. Еще 1,5 молекулы АТФ синтезируется из каждых 70 НАД+ и Н+. Это дает к 34 еще 105 молекул АТФ (т.е. всего 139) при окислении водорода, отщепляющегося при окислении каждой молекулы стеариновой кислоты.
Дополнительно 9 молекул АТФ образуются в цикле лимонной кислоты (помимо АТФ, получаемой при окислении водорода), по 1 на каждую из 9 молекул метаболизирующегося ацетил-КоА. Итак, при полном окислении 1 молекулы стеариновой кислоты образуются в сумме 148 молекул АТФ. С учетом того, что при взаимодействии стеариновой кислоты с КоА на начальной стадии метаболизма этой жирной кислоты расходуются 2 молекулы АТФ, чистый выход АТФ составляет 146 молекул.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
🎥 Видео
Расчет энергетической ценности и коэффициента Р/О при бета-окислении жирных кислотСкачать

Энергия окисления жирных кислот. БиохимияСкачать

Мобилизация жиров и регуляция. Бета окисление жирных кислот и регуляцияСкачать
Лекция 9.2. Окисление жирных кислотСкачать

Синтез и бета-окисление жирных кислотСкачать

Биохимия. Лекция 57. Липиды. Обмен триацилглицеролов и фосфолипидов. Окисление жирных кислотСкачать

ЛИПИДЫ 3: МОБИЛИЗАЦИЯ ЖИРОВ, БЕТА-ОКИСЛЕНИЕ ЖК, КЕТОНОВЫЕ ТЕЛА, ЭЙКОЗАНОИДЫСкачать

Метаболизм жиров. Окисление жирных кислот в митохондриях. Бета-оксидация и биосинтез нового жира.Скачать

Реакция на результаты ЕГЭ 2022 по русскому языкуСкачать

Нарушение бета-окисления жирных кислотСкачать

Окисление жирных кислотСкачать

Синтез жирных кислот и его регуляция. Влияние избыточного потребления углеводов на их синтезСкачать

Окислительно-восстановительные реакции в кислой среде. Продвинутый подход.Скачать

Биохимия | Синтез жиров и фосфолипидовСкачать

Лекция 9.4. Синтез ВЖКСкачать

ОБМЕН ЛИПИДОВ 2. Синтез высших жирных кислот.Скачать

Жирные кислоты - расшифровка анализа.Скачать
